
T-rex para niños
Datos para niños
Tyrannosaurus rex |
||
|---|---|---|
| Rango temporal: 68 Ma - 66 Ma Cretácico Superior (Maastrichtiense) | ||
 Reconstrucción en vida de un T. rex.
|
||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| (sin rango) | Opimoda Podiata Amorphea Opisthokonta Holozoa Filozoa Apoikozoa |
|
| Reino: | Animalia | |
| Subreino: | Eumetazoa | |
| (sin rango) | Bilateria | |
| Superfilo: | Deuterostomia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Infrafilo: | Gnathostomata | |
| Superclase: | Tetrapoda | |
| Clase: | Sauropsida | |
| Subclase: | Diapsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Coelurosauria | |
| Superfamilia: | Tyrannosauroidea | |
| Familia: | Tyrannosauridae | |
| Subfamilia: | Tyrannosaurinae | |
| Tribu: | Tyrannosaurini | |
| Género: | Tyrannosaurus Osborn, 1905 |
|
| Especie: | T. rex Osborn, 1905 |
|
| Sinonimia | ||
El Tyrannosaurus rex (que significa "lagarto tirano rey") es la única especie conocida del género fósil Tyrannosaurus. Fue un dinosaurio terópodo tiranosáurido que vivió al final del período Cretácico, hace unos 68 a 66 millones de años, en lo que hoy es América del Norte. Fue uno de los últimos dinosaurios no avianos antes de la extinción masiva del Cretácico-Terciario. También se le conoce como T. rex o simplemente tiranosaurio.
Como otros tiranosáuridos, el T. rex caminaba sobre dos patas (era bípedo). Tenía un cráneo enorme que se equilibraba con una cola larga y pesada. Sus patas traseras eran largas y fuertes, mientras que sus patas delanteras eran pequeñas, pero sorprendentemente robustas para su tamaño, y terminaban en dos dedos con garras. El Tyrannosaurus rex fue el tiranosáurido más grande conocido y uno de los mayores depredadores terrestres. Medía entre 12 y 13 metros de largo, 4 metros de alto hasta las caderas, y pesaba entre 6 y 9 toneladas.
Se han identificado más de 30 esqueletos de Tyrannosaurus rex, algunos casi completos. Se han encontrado tejidos y proteínas en al menos uno de estos fósiles. La gran cantidad de fósiles ha permitido a los científicos estudiar muchos aspectos de su biología, como su ciclo de vida y cómo se movía.
Contenido
- ¿Qué es el Tyrannosaurus Rex?
- Descubrimiento e investigación del T. rex
- Clasificación del T. rex
- ¿Cómo vivía el T. rex?
- ¿Dónde vivía el T. rex?
- El T. rex en la cultura popular
- Galería de imágenes
- Véase también
¿Qué es el Tyrannosaurus Rex?
El Tyrannosaurus rex tenía el cuerpo típico de su familia: una cabeza grande, un cuello corto en forma de "S", el cuerpo paralelo al suelo, patas delanteras pequeñas, patas traseras muy desarrolladas y una cola larga. Los ejemplares encontrados medían entre 11.1 y 13 metros de longitud y pesaban entre 5.6 y 9.5 toneladas.
Su cráneo era grande, de hasta 1.4 metros, con aberturas para los ojos y la nariz. Tenía muchos huesos fusionados, lo que le daba una estructura muy fuerte, algo inusual en otros terópodos que solían tener huesos más ligeros. El cuello era corto, grueso y musculoso para sostener su enorme cabeza.
La cola era pesada y larga, con más de cuarenta vértebras, para equilibrar su gran cuerpo y cabeza. Para que el animal no fuera demasiado pesado, muchos de sus huesos eran huecos, lo que reducía su peso sin perder fuerza.
¿Qué tan grande era el T. rex?


El esqueleto casi completo más grande, llamado Sue (FMNH PR2081), medía entre 12.3 y 12.4 metros de largo y entre 3.66 y 3.96 metros de alto hasta las caderas. Su peso se ha estimado entre 4.5 y 8.4 toneladas. Otro espécimen, Scotty (RSM P2523.8), se cree que medía 13 metros de largo y pesaba unas 8.87 toneladas métricas, siendo el más grande conocido.
Aunque era más largo que el terópodo del Jurásico Allosaurus y similar en tamaño al Carcharodontosaurus africano, el Tyrannosaurus era más pequeño que otros terópodos del Cretácico como el Spinosaurus y el Giganotosaurus.
¿Cómo era el cráneo del T. rex?

El cráneo más grande de Tyrannosaurus rex medía 1.52 metros de largo. Tenía grandes aberturas, llamadas fenestras, que reducían su peso y daban espacio para los músculos. El cráneo del Tyrannosaurus era muy ancho en la parte trasera y estrecho en el hocico, lo que le permitía tener una visión binocular (ver con ambos ojos a la vez) muy buena.
Los huesos del cráneo eran muy fuertes y algunos estaban fusionados, lo que lo hacía más resistente. Esto ayudaba al Tyrannosaurus a tener una mordida extremadamente potente, mucho más fuerte que la de otros carnívoros. La parte delantera de su mandíbula superior tenía forma de "U", lo que le permitía arrancar más carne y hueso con cada mordida.
La mandíbula inferior era robusta y tenía trece dientes en la parte delantera. Las mandíbulas tenían muchos pequeños agujeros que podrían haber sido para un sistema sensorial, similar al de los cocodrilos, o para estructuras como escamas o labios.
Los dientes del T. rex

Los dientes del Tyrannosaurus rex eran de diferentes formas. Los dientes de la parte delantera de la mandíbula superior estaban muy juntos, tenían forma de "D" en su sección transversal y estaban curvados hacia atrás. Estas características ayudaban a que los dientes no se rompieran al morder y desgarrar.
El resto de los dientes eran robustos, parecidos a "plátanos afilados", y estaban más separados. Los dientes de la mandíbula superior eran más grandes que los de la parte trasera de la mandíbula inferior. El diente más grande de Tyrannosaurus encontrado hasta ahora medía unos 30.5 centímetros de largo, incluyendo la raíz, siendo el diente más grande de cualquier dinosaurio carnívoro descubierto.
Esqueleto y extremidades del T. rex

La columna vertebral del Tyrannosaurus tenía 10 vértebras en el cuello, 13 en la espalda y 5 en el sacro. El número de vértebras en la cola variaba, pero probablemente eran al menos 40. El cuello era corto y musculoso para sostener su enorme cabeza.
Las patas traseras eran muy largas en proporción a su cuerpo. En el pie, el tercer hueso del metatarso (el pie) estaba apretado entre el segundo y el cuarto, formando una unidad fuerte que le ayudaba a moverse de forma eficiente.

Las patas delanteras del Tyrannosaurus eran pequeñas en comparación con el resto de su cuerpo, midiendo solo 1 metro de largo. Sin embargo, no eran inútiles. Los huesos de sus brazos tenían grandes áreas para la inserción de músculos, lo que indica que eran sorprendentemente fuertes. Tenían solo dos dedos con garras.
La función de estas pequeñas patas delanteras ha sido muy debatida. Algunos científicos sugieren que podrían haber ayudado al animal a levantarse del suelo o a sujetar a la presa mientras sus poderosas mandíbulas la mataban. Un estudio biomecánico mostró que el músculo bíceps de un Tyrannosaurus rex adulto podía levantar 199 kilgramos por sí solo.
Descubrimiento e investigación del T. rex
Hasta 2006, se habían encontrado 30 especímenes de Tyrannosaurus rex, incluyendo solo tres cráneos completos. Todos los fósiles se han hallado en Norteamérica.
Primeros hallazgos del T. rex
Los primeros dientes que hoy se reconocen como de Tyrannosaurus rex fueron encontrados en 1874 por A. Lakes en Golden (Colorado)Golden, Colorado. A principios de la década de 1890, J. B. Hatcher recolectó huesos en el este de Wyoming.
En 1892, Edward Drinker Cope encontró dos fragmentos de vértebras de un gran dinosaurio y los llamó Manospondylus gigas. Más tarde, en 1912, Henry Fairfield Osborn los atribuyó a Tyrannosaurus rex. Aunque Manospondylus gigas fue nombrado primero, las reglas de nomenclatura científica permiten que Tyrannosaurus rex siga siendo el nombre válido debido a su uso extendido.

Henry Fairfield Osborn, presidente del Museo Americano de Historia Natural, describió oficialmente al Tyrannosaurus rex en 1905. El nombre Tyrannosaurus viene del griego "tirano" y "lagarto", y rex es la palabra latina para "rey". Así, Tyrannosaurus rex significa "el rey de los lagartos tiranos", destacando su gran tamaño y su dominio.
El trabajo de Barnum Brown
Barnum Brown, un explorador del Museo Americano de Historia Natural, encontró el segundo esqueleto de T. rex en Wyoming en 1900. Luego encontró otro esqueleto parcial en Montana en 1902. El primer espécimen fue llamado originalmente Dynamosaurus imperiosus, pero en 1906, Osborn reconoció que ambos esqueletos eran de la misma especie y eligió Tyrannosaurus como el nombre oficial.
En total, Barnum Brown encontró cinco esqueletos parciales de T. rex. Su cuarto hallazgo, el más importante, está expuesto en el Museo Americano de Historia Natural en Nueva York.
Descubrimientos importantes desde 1960
A partir de la década de 1960, el interés en el Tyrannosaurus resurgió, llevando al descubrimiento de 42 esqueletos más en el oeste de Norteamérica. La década de 1990 fue especialmente fructífera, con el hallazgo de dos de los esqueletos más completos: Sue y Stan.

El esqueleto apodado Stan, encontrado en 1987 en Dakota del Sur, está completo en un 65%. Se exhibe en el Museo Black Hills de Historia Natural. Este Tyrannosaurus (BHI 3033) tiene muchas marcas de lesiones curadas en sus huesos, incluyendo fracturas en costillas y cuello.

Sue, el esqueleto fósil de T. rex más completo (más del 85%) y grande conocido hasta entonces, fue descubierto por Susan Hendrickson en 1990 en Dakota del Sur. Después de una disputa legal, el fósil fue vendido en subasta por 7.6 millones de dólares y ahora se exhibe en el Museo Field de Historia Natural en Chicago. El estudio de los huesos de Sue mostró que alcanzó su tamaño completo a los 19 años y vivió hasta los 28 años.
En 2001, se descubrió el 50% del esqueleto de un Tyrannosaurus juvenil, apodado Jane, en Montana. Este es el ejemplar juvenil más completo y mejor conservado hasta la fecha y se exhibe en el Museo Burpee de Historia Natural en Rockford, Illinois.
En 2005, se anunció el hallazgo de tejido blando dentro de un hueso de la pata de un T. rex de hace 68 millones de años. Se encontraron vasos sanguíneos flexibles y tejido elástico. Si este material es original, podría ayudar a entender el ADN de los dinosaurios.
En 2006, la Universidad Estatal de Montana reveló que tenía el cráneo de Tyrannosaurus más grande encontrado hasta ahora, el MOR-008, que mide 149.9 centímetros de largo.
Un estudio de 2021 estimó que el número total de T. rex que vivieron en el planeta fue de unos 2.500 millones, a lo largo de 127.000 generaciones.
¿Había otras especies de Tyrannosaurus?

En 1955, se nombró a Tyrannosaurus bataar en Mongolia, pero en 1965 se renombró como Tarbosaurus bataar. Aunque algunos científicos lo consideran una segunda especie de Tyrannosaurus, la mayoría cree que Tarbosaurus es un género separado debido a diferencias en el cráneo y la forma en que se distribuían las fuerzas al morder.
Otros fósiles de tiranosáuridos encontrados en las mismas zonas que Tyrannosaurus rex se han considerado originalmente especies diferentes, pero ahora se cree que son ejemplares jóvenes de Tyrannosaurus rex.
¿Qué es el Nanotyrannus?

Un cráneo pequeño pero muy completo encontrado en Montana fue clasificado como una nueva especie, Nanotyrannus, que significa "tirano enano". Se estimó que medía unos 5.2 metros de largo. Sin embargo, en 1999, un análisis detallado sugirió que era un Tyrannosaurus rex juvenil.
Las opiniones sobre si Nanotyrannus es una especie válida o un T. rex joven están divididas. Algunos científicos señalan pequeñas diferencias, como un mayor número de dientes en Nanotyrannus. Sin embargo, otros estudios indican que estas diferencias pueden ser variaciones normales durante el crecimiento de un T. rex joven.
En 2020, un estudio de Holly Woodward y sus colegas concluyó que los especímenes atribuidos a Nanotyrannus eran inmaduros y probablemente pertenecían a T. rex.
¿Podría haber más especies de T. rex?

En un estudio de 2022, Gregory S. Paul y sus colegas propusieron que el Tyrannosaurus rex tradicionalmente conocido podría ser en realidad tres especies diferentes: Tyrannosaurus rex, Tyrannosaurus imperator (emperador) y Tyrannosaurus regina (reina). Esta idea se basó en las variaciones en el tamaño y la robustez de los huesos del fémur y otras partes del esqueleto.
Sin embargo, muchos otros paleontólogos han criticado este estudio, argumentando que las diferencias observadas son parte de la variación natural dentro de una sola especie. La discusión sobre si el Tyrannosaurus tiene una o varias especies sigue abierta.
Clasificación del T. rex
El Tyrannosaurus es el género principal de la superfamilia Tyrannosauroidea, la familia Tyrannosauridae y la subfamilia Tyrannosaurinae. Los tiranosáuridos se consideran parte de los celurosaurios, un grupo de dinosaurios generalmente más pequeños.
Muchos análisis genéticos han encontrado que el Tarbosaurus bataar es el pariente más cercano del T. rex. El descubrimiento del tiranosáurido Lythronax también sugiere que Tarbosaurus y Tyrannosaurus están estrechamente relacionados.
¿Cómo vivía el T. rex?
Cerebro y sentidos del T. rex
Un estudio de Lawrence Witmer y Ryan Ridgely encontró que el Tyrannosaurus tenía sentidos muy desarrollados. Podía mover sus ojos y cabeza rápidamente, detectar sonidos de baja frecuencia (lo que le permitía rastrear presas a distancia) y tenía un sentido del olfato muy agudo.

Se cree que el Tyrannosaurus tenía una visión muy buena, incluso mejor que la de los halcones modernos. Podía ver objetos a una distancia de hasta 6000 metros. Su gran sentido del olfato, comparable al de los buitres, le permitía detectar cadáveres a grandes distancias.
El T. rex también tenía una cóclea (parte del oído interno) muy larga, lo que sugiere que la audición era un sentido importante para él, especialmente para escuchar sonidos de baja frecuencia. Un estudio de 2017 sugirió que el hocico de los tiranosáuridos era muy sensible, lo que podría haberles ayudado a sentir la temperatura de sus nidos o a manipular suavemente los huevos y las crías, como hacen los cocodrilos modernos.
El cerebro del Tyrannosaurus era relativamente grande en comparación con otros dinosaurios no avianos, aunque su tamaño seguía estando dentro del rango de los reptiles modernos.
Postura del T. rex

El Tyrannosaurus, como todos los terópodos, caminaba sobre dos patas. Sus patas tenían un tejido acolchado que actuaba como un resorte. En los siglos XIX y XX, se pensaba que el T. rex se paraba muy erguido, como un canguro, con la cola arrastrando por el suelo. Esta idea se reforzó cuando el primer esqueleto completo de Tyrannosaurus rex se exhibió en 1915 en esta posición.
Sin embargo, en la década de 1970, los científicos se dieron cuenta de que esta postura era incorrecta. Un animal vivo no podría haberla mantenido sin dislocarse varias articulaciones. Las representaciones modernas del T. rex en museos y películas, como Parque Jurásico, muestran su cuerpo más paralelo al suelo y la cola extendida hacia atrás para equilibrar la cabeza.

Para sentarse, el Tyrannosaurus pudo haber apoyado su peso en la parte trasera de su pelvis. Levantarse de nuevo podría haber implicado el uso de sus pequeñas patas delanteras para estabilizarse.
Crecimiento del T. rex

El estudio de varios ejemplares jóvenes de Tyrannosaurus ha permitido a los científicos entender cómo crecían y cuánto tiempo vivían. El ejemplar más pequeño conocido pesaba solo 30 kilogramos y tenía dos años al morir, mientras que Sue, el más grande, tenía 28 años, lo que podría ser la edad máxima de la especie.
El Tyrannosaurus rex crecía lentamente hasta los 14 años, cuando su tamaño aumentaba drásticamente. Durante esta fase, ganaba unos 600 kilogramos al año durante cuatro años. A los 18 años, su crecimiento se ralentizaba mucho. Este cambio repentino en el crecimiento podría indicar que alcanzaban la madurez física.
Se ha encontrado tejido medular (un tipo de tejido óseo que solo se encuentra en hembras de aves durante la ovulación) en el fémur de un Tyrannosaurus de 16 a 20 años, lo que sugiere que era una hembra en edad reproductiva.
Piel y plumas del T. rex

No hay pruebas directas de que el T. rex tuviera plumas. Sin embargo, muchos de sus parientes cercanos sí las tenían, lo que sugiere que el T. rex también podría haberlas tenido. Por ejemplo, el tiranosauroide primitivo Dilong paradoxus tenía filamentos de protoplumas (plumas primitivas) en la cola.
Sin embargo, algunas impresiones de piel de tiranosaurios adultos muestran escamas, como las de otros dinosaurios. Se ha propuesto que la presencia de plumas o escamas podría depender del tamaño del animal o del clima. En climas fríos, las plumas habrían servido de aislamiento térmico, pero en climas cálidos, un animal tan grande podría haberse sobrecalentado con una cubierta de plumas.
El descubrimiento del gigante Yutyrannus huali, que tenía plumas en gran parte de su cuerpo, puso en duda la idea de que las plumas solo se encontraban en dinosaurios pequeños. Un estudio de 2017 sugirió que si el Tyrannosaurus tenía plumas, estas se limitarían a la parte superior de su cuerpo.
¿Cómo regulaba su temperatura el T. rex?
No está claro si los tiranosaurios eran de "sangre fría" (ectotérmicos) o de "sangre caliente" (endotérmicos). Hasta los años 1960, se pensaba que la mayoría de los dinosaurios eran de "sangre fría". Sin embargo, científicos como Robert T. Bakker y John Ostrom argumentaron que el Tyrannosaurus debió haber sido de "sangre caliente" para tener un estilo de vida tan activo.
Las altas tasas de crecimiento de los Tyrannosaurus rex jóvenes, similares a las de mamíferos y aves, apoyan la idea de un metabolismo alto. Un estudio de los huesos de Tyrannosaurus encontró que la diferencia de temperatura entre el centro del cuerpo y las extremidades era pequeña, lo que sugiere que mantenían una temperatura corporal constante, como los mamíferos.
¿Qué tan rápido se movía el T. rex?
.JPG)
El Tyrannosaurus tenía patas largas, pero hay desacuerdo sobre su velocidad. Algunos científicos creen que los animales pesados con patas como pilares no pueden correr muy rápido. Las estimaciones de velocidad varían desde 5 km/h hasta 72 km/h, aunque esta última es muy poco probable.
La mayoría de las investigaciones recientes sugieren que el Tyrannosaurus no podía correr a más de 40 km/h. Un estudio de 2002 indicó que para correr a más de 40 km/h, el T. rex necesitaría músculos en las piernas que representaran más del 40-86% de su masa corporal total, lo cual es inviable. Es más probable que caminara o trotara a velocidades de unos 18 km/h.
Las caídas eran muy peligrosas para el T. rex debido a su gran tamaño. Si se caía mientras se movía rápidamente, podría sufrir lesiones graves o incluso mortales. Sin embargo, algunos argumentan que, al igual que las jirafas, el Tyrannosaurus podría haber asumido estos riesgos cuando era necesario.
Un estudio de 2019 sugirió que el Tyrannosaurus y otros tiranosáuridos eran más ágiles que otros terópodos de tamaño similar, lo que les permitía hacer giros rápidos y pivotar su cuerpo al cazar. Otro estudio de 2020 indicó que eran caminantes muy eficientes, lo que les permitía ahorrar energía al buscar alimento.
Huellas del T. rex

Es difícil atribuir una huella específica a un Tyrannosaurus porque los pies de los diferentes terópodos dejaban huellas muy similares de tres dedos. Sin embargo, se han asignado provisionalmente dos huellas fósiles aisladas al Tyrannosaurus.
La primera fue descubierta en Nuevo México en 1983 y se llamó Tyrannosauripus pillmorei. Medía 83 centímetros de largo por 71 de ancho. Una segunda huella, de 72 por 76 centímetros, fue descubierta en Montana en 2007. Aunque no es seguro, el Tyrannosaurus es el único terópodo grande conocido en esa formación.
¿Cómo se comportaba el T. rex en grupo?

]]
Algunos científicos, como Philip J. Currie, han sugerido que el Tyrannosaurus pudo haber cazado en manadas, basándose en el hallazgo de varios esqueletos de T. rex muy cerca unos de otros en Dakota del Sur. La caza cooperativa podría haber sido una estrategia efectiva para cazar presas grandes y peligrosas como el Triceratops y el Ankylosaurus.
Sin embargo, otros investigadores han criticado esta hipótesis, señalando que la evidencia de comportamiento en grupo en otros tiranosáuridos es débil y podría explicarse por eventos como sequías o inundaciones que hicieron que los dinosaurios murieran juntos. También se ha sugerido que, en lugar de cazar en manada, podrían haber tenido un comportamiento similar al de los dragones de Komodo, donde varios individuos se acercan a un cadáver.
Se ha encontrado evidencia de ataques entre Tyrannosaurus jóvenes. El cráneo de Jane, un Tyrannosaurus juvenil, muestra heridas curadas en la mandíbula superior y el hocico, lo que sugiere que fue atacado por otro Tyrannosaurus joven.
¿Qué comía el T. rex?
.jpg)
Las grandes mandíbulas del Tyrannosaurus medían más de 1 metro y estaban llenas de dientes afilados y robustos de hasta 19 centímetros. Se ha calculado que su fuerza de mordida era la más poderosa de cualquier animal terrestre, capaz de ejercer una presión de 3.6 a 5.8 toneladas. Esto le permitía aplastar huesos y consumir completamente los cadáveres de grandes dinosaurios.
La mayoría de los paleontólogos creen que el Tyrannosaurus era tanto un depredador activo como un carroñero, como la mayoría de los grandes carnívoros modernos. Era el carnívoro más grande de su entorno y probablemente cazaba hadrosáuridos, ceratópsidos y anquilosaurianos.
¿Era el T. rex un cazador o un carroñero?
El debate sobre si el Tyrannosaurus era un depredador o un carroñero puro es muy antiguo. Jack Horner, un experto en hadrosáuridos, es el principal defensor de la idea de que el Tyrannosaurus era exclusivamente carroñero. Sus argumentos incluyen:
- Las patas delanteras del Tyrannosaurus eran cortas y no tenían suficiente fuerza para sujetar a la presa.
- Tenía un sentido del olfato muy desarrollado, lo que le permitiría detectar cadáveres a grandes distancias, como los buitres.
- Sus dientes podían machacar huesos, lo que le permitiría extraer el máximo de alimento de los restos de un animal.
- Si el Tyrannosaurus caminaba en lugar de correr, esto sugeriría que era carroñero, ya que algunas de sus presas potenciales eran rápidas.

Sin embargo, otros indicios sugieren que el Tyrannosaurus era un cazador:
- Sus ojos estaban orientados hacia adelante, dándole una visión binocular mejor que la de los halcones modernos, una característica común en los depredadores.
- Se han encontrado huesos de Edmontosaurus con marcas de mordeduras de Tyrannosaurus que luego sanaron, lo que demuestra que el Edmontosaurus sobrevivió a un ataque, indicando que el Tyrannosaurus intentó cazarlo activamente.
- También hay evidencia de interacción agresiva con Triceratops, con marcas de dientes de Tyrannosaurus en sus huesos.
La mayoría de los paleontólogos aceptan que el Tyrannosaurus era tanto un depredador activo como un carroñero, como la mayoría de los grandes carnívoros modernos. Los carnívoros actuales rara vez son solo cazadores o solo carroñeros; su comportamiento depende de la disponibilidad de alimento. Si el Tyrannosaurus robaba presas a depredadores más pequeños, su gran tamaño habría sido suficiente para ahuyentarlos.
¿Saliva infecciosa?
Se ha sugerido que la saliva del Tyrannosaurus podría haber sido infecciosa para sus presas, similar al dragón de Komodo. Esta idea se basa en que los espacios entre los dientes aserrados podrían haber retenido fibras de carne en putrefacción con bacterias. Sin embargo, otros científicos han señalado diferencias en la forma de los dientes que hacen esta hipótesis menos probable.
Cuidado parental del T. rex
No hay pruebas directas de que el Tyrannosaurus cuidara a sus crías. Sin embargo, la escasez de fósiles de tiranosaurios jóvenes y nidos ha llevado a los investigadores a especular. Algunos sugieren que, al igual que sus parientes vivos más cercanos (aves y cocodrilos), el Tyrannosaurus pudo haber protegido y alimentado a sus crías.
¿Dónde vivía el T. rex?

El Tyrannosaurus vivía en todo el oeste de Norteamérica, desde Alberta (Canadá) hasta Coahuila en México, justo antes de la extinción de los dinosaurios. Solía habitar en llanuras inundables y bosques subtropicales, acechando a sus presas en zonas con ríos, lagos y vegetación exuberante, incluyendo cicadáceas, helechos, plantas florecidas y árboles como coníferas y araucarias.

En la época del T. rex, Norteamérica tenía un paisaje con elementos familiares y extraños. Había tortugas de cuero, cocodrilos, lucios y peces pipa similares a los actuales. También ranas y lagartos varanos. Las plantas dominantes eran helechos, palmas, magnolias y álamos. Las coníferas como las secuoyas eran comunes.
Otros habitantes del paisaje eran los pterosaurios gigantes como el Quetzalcoatlus, que volaban con alas de más de 12 metros. Otros terópodos más pequeños, como los dromeosáuridos, medían menos de 5 metros. Manadas de Triceratops y Edmontosaurus vagaban por la tierra. Los mamíferos eran pequeños y nocturnos, parecidos a ratas y musarañas.
Formaciones donde se encontraron fósiles
Se han encontrado muchos restos de Tyrannosaurus en la Formación Hell Creek. Durante el Maastrichtiense, esta zona era subtropical, con un clima cálido y húmedo. El Tyrannosaurus compartía este ecosistema con otros dinosaurios como el Triceratops, el Edmontosaurus y el Ankylosaurus.
Otra formación con restos de Tyrannosaurus es la Formación Lance de Wyoming, que se cree que era un ambiente pantanoso. La fauna era muy similar a la de Hell Creek.
En el sur de su área de distribución, el Tyrannosaurus vivía junto al saurópodo Alamosaurus y otros ceratópsidos y hadrosaurios.
¿Cuántos T. rex existieron?

Según estudios de 2021, se estima que en un momento dado, la población de Tyrannosaurus adultos era de unos 20,000 individuos. A lo largo de la existencia del género, se calcula que vivieron aproximadamente 2,500 millones de Tyrannosaurus en total hasta su extinción.
El T. rex en la cultura popular
Desde que fue descrito en 1905, el Tyrannosaurus rex se ha convertido en el dinosaurio más famoso en la cultura popular. Es el único dinosaurio conocido comúnmente por su nombre científico completo (Tyrannosaurus rex), y su abreviatura T. rex también es muy usada.
Galería de imágenes
-
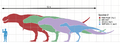
Varios especímenes de Tyrannosaurus rex en comparación con un humano.
-
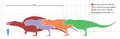
Comparación del tamaño de algunos dinosaurios terópodos gigantes y un ser humano.
-

Perfil del cráneo de Tyrannosaurus AMNH 5027.
-

Dos dientes de la mandíbula inferior del ejemplar MOR 1125, B-rex, mostrando la variación en tamaño de los dientes en un mismo individuo.
-

Fúrcula del espécimen Sue
-

Miembro anterior derecho de Tyrannosaurus
-

Restauración del esqueleto por William D. Matthew de 1905, una de las primeras reconstrucciones de Tyrannosaurus rex publicadas.
-

Cráneo tipo de T. rex, del Museo Carnegie de Historia Natural. Reconstruido en forma incorrecta tomando como modelo uno de Allosaurus.
-

Tyrannosaurus rex, réplica del espécimen BHI 3033, o Stan en el Real Instituto Belga de Ciencias Naturales en Bruselas, Bélgica
-

Sue, Tyrannosaurus del Field Museum, Chicago.
-

Fémur del espécimen MOR 1125 de T. rex, del cual se obtuvieron la matriz desmineralizada y péptidos (en los recuadros).
-

Diagrama que muestra las diferencias entre los cráneos de Tarbosaurus (A) y de Tyrannosaurus (B).
-

Cráneo holotipo de Nanotyrannus lancensis, posible ejemplar juvenil de Tyrannosaurus.
-

Modelo de Tyrannosaurus rex basado en los últimos hallazgos, de que poseía una complexión robusta con protoplumas.
-

Tyrannosaurus rex
-

Tarbosaurus bataar
-

Molde de la caja craneana en el Museo Australiano, Sídney.
-

Recreación de un Tyrannosaurus rex basado en los últimos estudios, con la postura correcta
-

T. rex podría haber usado sus patas delanteras para levantarse después de haber estado en una postura de descanso, como se ve aquí
-
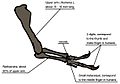
Diagrama que ilustra la anatomía del brazo
-
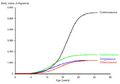
Curva de crecimiento de T. rex comparada con la de otros tiranosáuridos. Basada en Erickson et al. (2004).
-
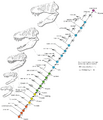
Diagrama que muestra las etapas de crecimiento
-

Recreación de Tyrannosaurus rex con plumas, basado en el espécimen AMNH 5027.
-

Restauración de un individuo (basado en MOR 980) con infecciones parasitarias
-

Antigua representación de T. rex (con una postura incorrecta, ver más abajo) en su hábitat natural. Dibujo de Charles R. Knight.
-

Tyrannosaurus (izquierda), y otros animales de la Formación Hell Creek
-

La pata derecha de T. rex (lateral) fotografiada en el Museo de Historia Natural de la Universidad de Oxford.
-

Un ave de seis toneladas habría necesitado músculos de las piernas que constituyeran casi el 100 % de su masa corporal para correr. Siendo realistas, T. rex tenía los músculos para correr a unos 18 km/h.
-

Posible huella en Nuevo México. Contramolde en relieve invertido (localizado en la cara inferior de un estrato).
-

Mandíbula de tiranosaurio.
-

Un Allosaurus devorando carroña de un saurópodo. Dibujo de Charles R. Knight.
-

Esqueletos montados de diferentes grupos de edad, Museo de Historia Natural del Condado de Los Ángeles]]
-

Gráfico del censo promediado en el tiempo para dinosaurios de cuerpo grande de toda la Formación Hell Creek en el área de estudio.


.jpg)





Véase también
 En inglés: Tyrannosaurus Facts for Kids
En inglés: Tyrannosaurus Facts for Kids
- Anexo: Géneros válidos de dinosaurios
- Tyrannosauridae
- Especímenes de Tyrannosaurus rex
