
Cretácico para niños
| Era Eratema |
Período Sistema |
Inicio (millones años) |
|---|---|---|
| Mesozoica | Cretácico | 143,1 |
| Jurásico | 201,4 ±0,2 | |
| Triásico | 251,902 ±0,024 |

El Cretácico es un período muy importante en la historia de la Tierra. Es el tercer y último período de la Era Mesozoica, también conocida como la "Era de los Dinosaurios". El Cretácico comenzó hace unos 143,1 millones de años y terminó hace 66 millones de años. Duró aproximadamente 77 millones de años, lo que lo convierte en el período más largo de la Era Fanerozoica.
Su nombre viene de la palabra en latín creta, que significa "tiza". Esto se debe a que en este período se formaron grandes depósitos de tiza, una roca blanda hecha de restos de pequeños organismos marinos. Un geólogo belga llamado Jean d'Omalius d'Halloy fue quien lo nombró así en 1822.
El Cretácico se divide en dos partes principales: el Cretácico inferior (la primera parte) y el Cretácico superior (la última parte).
Durante este tiempo, la vida en los océanos y en la tierra era una mezcla de formas antiguas y nuevas. Al final del Cretácico, hubo un gran evento que cambió el planeta para siempre. Se cree que un meteorito gigante chocó contra la Tierra en lo que hoy es la península de Yucatán en México. Este impacto pudo haber causado una extinción masiva que marcó el fin de los dinosaurios y de la Era Mesozoica.
A mediados del Cretácico, se formó más de la mitad del petróleo que conocemos hoy en día. Grandes reservas se encuentran en lugares como el golfo Pérsico y la región entre el golfo de México y la costa de Venezuela.
Contenido
¿Cómo se divide el Cretácico?
El Cretácico se organiza en diferentes etapas, llamadas edades o pisos. Aquí te mostramos cómo se dividen, desde las más recientes hasta las más antiguas:
| Era Eratema |
Periodo Sistema |
Época Serie |
Edad Piso |
Inicio, en millones de años |
|---|---|---|---|---|
| Mesozoico | Cretácico | Superior / Tardío | Maastrichtiense Maastrichtiano |
|
| Campaniense Campaniano |
||||
| Santoniense Santoniano |
||||
| Coniaciense Coniaciano |
||||
| Turoniense Turoniano |
||||
| Cenomaniense Cenomaniano |
||||
| Inferior / Temprano | Albiense Albiano |
|||
| Aptiense Aptiano |
121,4±0,6 | |||
| Barremiense Barremiano |
||||
| Hauteriviense Hauteriviano |
||||
| Valanginiense Valanginiano |
||||
| Berriasiense Berriasiano |
143,1 | |||
| Jurásico | 201,4±0,2 | |||
| Triásico | 251,9±0,024 | |||
¿Cómo era la Tierra en el Cretácico?
Los continentes y los océanos

Durante el Cretácico, el nivel del mar subió mucho, más que en cualquier otro momento de la historia de la Tierra. Esto hizo que muchas zonas que antes eran tierra firme se inundaran. En su punto más alto, solo el 18% de la superficie terrestre estaba por encima del agua.
El supercontinente Pangea se había estado separando durante la Era Mesozoica. Al principio del Cretácico, existían dos grandes continentes: Laurasia (que incluía lo que hoy es Norteamérica, Europa y Asia) y Gondwana (que incluía Sudamérica, África, la Antártida, Australia y la India). Estos continentes estaban separados por un gran océano llamado el mar de Tetis.
A medida que el Cretácico avanzaba, los continentes se movían y se parecían cada vez más a los que conocemos hoy. La India y Madagascar se separaron de África, y la Antártida y Australia se alejaron de Sudamérica. Estos movimientos crearon nuevos océanos, como el Océano Atlántico y el océano Índico.
En Norteamérica, una gran masa de agua se extendía desde el Polo Norte hasta México, dividiendo el continente. Todos estos cambios hicieron que la Tierra se dividiera en muchas masas de tierra separadas. Esto fue muy importante porque permitió que diferentes tipos de animales y plantas evolucionaran de forma única en cada lugar.
El clima del Cretácico

Las temperaturas en la Tierra subieron mucho durante el Cretácico, alcanzando su punto más alto hace unos 100 millones de años. En ese momento, casi no había hielo en los polos. Los océanos tropicales eran mucho más cálidos que ahora, y las temperaturas en las profundidades del océano también eran muy altas.
El clima era más suave en general, con menos diferencias de temperatura entre los polos y el ecuador. Esto hizo que las corrientes de aire y las corrientes oceánicas fueran más lentas, lo que llevó a que los océanos estuvieran más estancados en algunas zonas. Después de la mitad del Cretácico, las temperaturas comenzaron a bajar lentamente, y al final del período, el clima se volvió más frío.
¿Qué animales vivieron en el Cretácico?
El Cretácico fue la época de mayor esplendor para los dinosaurios, que vivían en todas las regiones del planeta. Al final del período, la mayoría de los animales, tanto terrestres como marinos, sufrieron la extinción masiva del K-Pg.
Animales marinos

En los océanos del Cretácico, el plancton (pequeños organismos que flotan en el agua) se parecía mucho al actual. Las diatomeas y los dinoflagelados, que son tipos de plancton, se expandieron mucho. También había muchos ammonites (moluscos con conchas en espiral) y belemnites (parecidos a los calamares).
Los ictiosaurios, que eran reptiles marinos parecidos a los delfines, casi desaparecieron al principio del Cretácico. Su lugar fue ocupado por nuevos tiburónes grandes y peces óseos gigantes como el Xiphactinus, que medía entre 2 y 4 metros. También había tortugas marinas enormes como Archelon, de más de 3 metros de largo.
Los plesiosaurios, reptiles marinos de cuello largo, podían medir hasta 12 metros. Pero los depredadores más temibles del Cretácico superior eran los mosasaurios. Estos lagartos gigantes, parientes de las serpientes actuales, podían alcanzar los 17 metros de longitud. Tenían mandíbulas largas con dientes afilados y se alimentaban de peces y ammonites.
-

Jeletzkytes (un ammonite)
-

Belemnitella (un belemnite)
-

Tusoteuthis (un cefalópodo)
-

Xiphactinus (un pez gigante)
-

Archelon (una tortuga marina gigante)
-
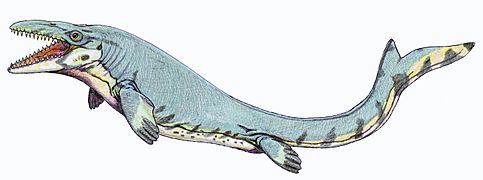
Mosasaurus (un reptil marino)
-

Libonectes (un plesiosaurio)








En el fondo del mar, los crustáceos como los cangrejos y las langostas se hicieron más comunes. Los bivalvos (como las almejas) desarrollaron conchas más gruesas o espinas para protegerse de los depredadores. Un grupo de bivalvos llamados rudistas formaban arrecifes enormes, casi tan grandes como los arrecifes de coral.
Animales aéreos
Los pterosaurios, reptiles voladores, se hicieron más grandes y perdieron sus dientes a lo largo del Cretácico. Uno de los más famosos fue el Pteranodon, con una gran cresta en la cabeza. Las aves fósiles son raras de encontrar en este período, pero se han descubierto algunas en China y España, como Iberomesornis y Concornis.
-

Ornithocheirus (un pterosaurio)
-

Pteranodon (un pterosaurio)
-

Ichthyornis (un ave)
-

Iberomesornis (un ave)




Los mamíferos existían, pero eran pequeños y no tan importantes como los dinosaurios. Los dinosaurios, especialmente los arcosaurios, tuvieron su mayor desarrollo en el Cretácico. Se han encontrado muchos fósiles de dinosaurios en lugares como Mongolia y el sureste de Inglaterra.
Los dinosaurios más conocidos del Cretácico
En esta época, los dinosaurios se diversificaron mucho, con muchas especies diferentes en tamaño y forma:
- Ornitópodos: Eran dinosaurios herbívoros que caminaban sobre dos o cuatro patas.
- En el Cretácico inferior, destacaron Iguanodon e Hypsilophodon. El Iguanodon era un bípedo de 10 metros de largo, con una boca parecida a la de un caballo y dientes que se reemplazaban. Tenía una púa afilada en el pulgar que quizás usaba como arma. El Hypsilophodon era más pequeño (3 a 5 metros) y muy rápido.
-
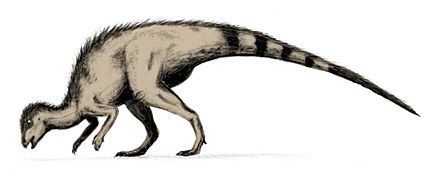
Hypsilophodon


-
- Los hadrosaurios (dinosaurios "pico de pato") eran grandes, de 10 a 15 metros. Tenían hocicos planos y cientos de dientes para masticar plantas. Muchos tenían crestas huecas en la cabeza, que usaban para hacer sonidos o para reconocerse entre ellos. Ejemplos famosos son Maiasaura y Parasaurolophus.
-
- Los ceratópsidos eran cuadrúpedos con cabezas grandes, picos y cuernos. También tenían un escudo óseo detrás de la cabeza para protegerse. El Triceratops era uno de los más grandes, con 9 metros de largo y cuernos de casi 1 metro. Otro ceratópsido conocido es Protoceratops.
-
- Los paquicefalosaurios eran ornitópodos bípedos con cráneos muy gruesos. Se cree que usaban sus cabezas para chocar entre sí, quizás para competir por parejas. El Pachycephalosaurus es un ejemplo.



-
- Los anquilosaurios eran cuadrúpedos blindados que se diversificaron en el Cretácico superior. Tenían placas óseas bajo la piel, cuernos y espinas. Algunos tenían una cola con una masa de hueso al final, como una maza, para defenderse.
- Terópodos: Eran dinosaurios carnívoros.
- El Baryonyx fue un carnívoro con una garra gigante en forma de hoz de 30 centímetros. Se cree que comía peces, ya que se encontraron escamas de pez en su esqueleto.
- El Tyrannosaurus rex (en Norteamérica) y el Tarbosaurus (en Mongolia) eran probablemente los depredadores más grandes de su tiempo. El Tyrannosaurus medía hasta 13 metros de largo y pesaba 5 toneladas, con dientes de 15 centímetros. Sus patas traseras eran fuertes, pero sus brazos eran muy cortos. Se cree que cazaba grandes herbívoros o se alimentaba de animales ya muertos.
-

Tarbosaurus








-
- Otros grandes carnívoros fueron Carnotaurus y Spinosaurus. También había terópodos más pequeños como Velociraptor, Deinonychus y Oviraptor. Se cree que muchos terópodos tenían plumas. Las aves son los únicos dinosaurios que sobrevivieron al final del Cretácico.
¿Qué plantas vivieron en el Cretácico?
La llegada de las angiospermas (plantas con flores) cambió mucho el paisaje de la Tierra. Al principio del Cretácico, las gimnospermas (plantas con semillas "desnudas", como los pinos) seguían siendo las dominantes. Las coníferas eran el grupo más exitoso.
Las angiospermas aparecieron en el Cretácico inferior, hace unos 130-120 millones de años. A diferencia de las gimnospermas, las angiospermas protegen sus semillas dentro de un ovario. Los fósiles más antiguos de angiospermas son granos de polen encontrados en Inglaterra.

En el Cretácico superior, ya existían unas 50 familias de plantas modernas, como hayas, higueras, robles y palmeras. El polen y las hojas de angiospermas se hicieron muy comunes, lo que demuestra su rápido crecimiento. Las primeras angiospermas crecían en áreas con mucha agua o inundaciones.
Las primeras flores e insectos

Las plantas con flores no aparecieron hasta el Cretácico, y las primeras no tenían pétalos. Su desarrollo fue posible gracias a su relación con los insectos. Se cree que insectos y plantas evolucionaron juntos. Los escarabajos y otros insectos pudieron haber polinizado las primeras plantas con estructuras parecidas a flores.
Se han encontrado muchos fósiles de insectos del Cretácico inferior, como libélulas, cucarachas, escarabajos y avispas. Algunos insectos tenían polen en su cuerpo, lo que indica que lo comían y ayudaban a polinizar.
La abeja fósil más antigua, Melittosphex burmensis, fue descubierta en 2006 en ámbar de Birmania. Tenía características de abeja y de avispa. La aparición de termitas en el Cretácico inferior y de abejas y hormigas en el superior, muestra avances importantes en el comportamiento social de los insectos.
La gran extinción al final del Cretácico

Al final del Cretácico, ocurrió un evento conocido como la extinción K-Pg (Cretácico-Paleógeno). Este evento causó la desaparición de los dinosaurios (excepto las aves), los pterosaurios, los reptiles nadadores (como plesiosaurios y mosasaurios), y muchos animales marinos como los ammonites.
Aunque fue una extinción importante, no fue la más grande de la historia de la Tierra. Muchos grupos de plantas y animales terrestres (insectos, caracoles, ranas, tortugas, lagartos, serpientes, cocodrilos y mamíferos) y la mayoría de los invertebrados marinos y peces lograron sobrevivir.

Se han propuesto varias ideas sobre qué causó esta extinción:
- Cambios climáticos: Algunos científicos creen que los cambios en las corrientes oceánicas y el descenso del nivel del mar causaron un enfriamiento global. Esto habría afectado a las faunas tropicales, mientras que las de latitudes más altas se mantuvieron.

- Impacto de un meteorito: Esta es la teoría más conocida. Se basa en el descubrimiento de una capa de iridio (un elemento raro en la Tierra, pero común en meteoritos) en rocas de esa época. La idea es que un meteorito gigante chocó con la Tierra, levantando una enorme nube de polvo que oscureció el cielo, bloqueó la luz del sol y causó un enfriamiento global. Esto habría afectado la cadena alimenticia, haciendo que muchas plantas y animales murieran.
Aunque hay pruebas de ambas ideas, el impacto del meteorito es la explicación más aceptada para la extinción repentina de muchos grupos, especialmente los dinosaurios.
Galería de imágenes
-

Estratos del límite Cretácico-Paleógeno en Alberta.
-
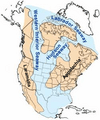
Mapa de los canales marinos de Norteamérica.


Véase también
 En inglés: Cretaceous Facts for Kids
En inglés: Cretaceous Facts for Kids
