
Cetacea para niños
Datos para niños Cetáceos |
||
|---|---|---|
| Rango temporal: 53,5 Ma - 0 Ma Eoceno - holoceno | ||
 Diferentes tipos de cetáceos.
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Clase: | Mammalia | |
| Subclase: | Theria | |
| Infraclase: | Placentalia | |
| Superorden: | Laurasiatheria Scrotifera |
|
| Orden: | Artiodactyla | |
| Suborden: | Whippomorpha | |
| Infraorden: | Cetacea Brisson, 1762 |
|
| Parvórdenes | ||
Los cetáceos (Cetacea) son un grupo de mamíferos placentarios que viven completamente en el agua. La palabra "cetáceo" viene del griego kētos, que significa 'ballena' o 'monstruo marino'. Fue usada por Aristóteles para referirse a los animales acuáticos que respiran con pulmónes.
Estos animales tienen un cuerpo con forma de huso, como el de los peces, lo que les ayuda a moverse muy bien en el agua. Sus patas delanteras se han convertido en aletas, y las traseras han desaparecido, aunque conservan algunos huesos pequeños dentro de su cuerpo. La aleta caudal (la cola) es horizontal y tiene dos partes.
Los cetáceos casi no tienen pelo y poseen una gruesa capa de grasa que los mantiene calientes. Existen unas ochenta especies de cetáceos, la mayoría vive en el mar, pero cinco especies de delfines habitan en agua dulce.
Los cetáceos que viven hoy se dividen en dos grupos: los misticetos (con barbas) y los odontocetos (con dientes). Un tercer grupo, los arqueocetos, ya no existe. Antes, los cetáceos se clasificaban como un grupo aparte, pero ahora se sabe que están relacionados con los hipopótamos y otros artiodáctilos, compartiendo antepasados de hace millones de años.
Entre los misticetos están las ballenas, que son los animales más grandes del mundo. La ballena azul es el animal más grande que ha existido en la Tierra, incluso más grande que los dinosaurios. Los odontocetos incluyen a los delfines y las orcas. La cetología es la ciencia que estudia a estos fascinantes animales.
Contenido
- ¿Cómo evolucionaron y se clasifican los cetáceos?
- ¿Cómo es el cuerpo de los cetáceos?
- ¿Cómo funcionan los cetáceos?
- ¿Cómo se reproducen los cetáceos?
- ¿Cómo se mantienen calientes?
- ¿Cómo obtienen agua?
- ¿Cuánto tiempo viven los cetáceos?
- ¿Dónde viven los cetáceos?
- ¿Cómo se comportan los cetáceos?
- ¿Cómo se relacionan con los humanos?
- ¿Cuál es el estado de conservación de los cetáceos?
- ¿Qué es la cetología?
- Galería de imágenes
- Véase también
¿Cómo evolucionaron y se clasifican los cetáceos?

La evolución de los cetáceos es muy interesante. Antes se pensaba que venían de unos animales terrestres parecidos a lobos con pezuñas. Sin embargo, estudios más recientes de biología molecular (analizando ADN) mostraron que los cetáceos están muy relacionados con los hipopotámidos (hipopótamos).
Esto significa que las ballenas y los delfines son parte del grupo de los artiodáctilos, que incluye a animales como ciervos, vacas y cerdos. Se cree que el primer antepasado de los cetáceos fue el Pakicetus, un artiodáctilo primitivo que vivió hace unos 50 millones de años. Este animal tenía dientes parecidos a los de las ballenas antiguas y una estructura especial en los huesos del oído.
Luego apareció el Ambulocetus, que fue el primer antepasado de los cetáceos en vivir tanto en tierra como en agua. Tenía patas más adaptadas para nadar. Más tarde, el basilosaurio, que vivió hace unos 38 millones de años, fue uno de los primeros cetáceos completamente acuáticos.
Clasificación de los cetáceos actuales
Los cetáceos que viven hoy se dividen en dos grandes grupos:
- Los misticetos (ballenas con barbas): Tienen unas estructuras en la parte superior de la boca, llamadas barbas, hechas de queratina. Estas barbas actúan como un filtro gigante para atrapar pequeños organismos como el plancton y el kril del agua.
* Incluyen familias como: Balaenidae (ballenas francas), Balaenopteridae (rorcuales), Eschrichtiidae (ballena gris) y Neobalaenidae (ballena pigmea).
- Los odontocetos (cetáceos con dientes): Se alimentan principalmente de peces y calamares. Tienen una habilidad especial llamada ecolocalización para "ver" su entorno usando sonidos.
* Incluyen familias como: Delphinidae (delfines oceánicos y orcas), Monodontidae (belugas y narvales), Phocoenidae (marsopas), Physeteridae (cachalotes), entre otras.
¿Cómo es el cuerpo de los cetáceos?

Los cetáceos, al adaptarse a la vida acuática, desarrollaron características especiales en su cuerpo:
- Su cuerpo es como un huso, muy liso y aerodinámico, similar al de un pez.
- Tienen una aleta en la espalda (aleta dorsal) hecha de tejido, que les ayuda a mantenerse estables al nadar.
- Sus patas delanteras se transformaron en aletas pectorales, que parecen remos.
- La cola es plana y tiene dos lóbulos, moviéndose de arriba abajo para impulsarse.
- No tienen patas traseras visibles, solo pequeños huesos escondidos.
- Respiran por un orificio en la parte superior de la cabeza, llamado espiráculo.
- No tienen pelo ni orejas externas.
- Sus partes externas están ocultas para que su cuerpo sea más liso.
Tamaño y forma del cuerpo
La forma del cuerpo de los cetáceos es muy parecida a la de los peces, lo que les permite moverse con agilidad en el agua. Los misticetos suelen ser más robustos, mientras que los odontocetos son más rápidos.
Los cetáceos incluyen algunos de los animales más grandes de la Tierra. La ballena azul puede medir hasta 30 metros de largo, ¡es el animal más grande de todos los tiempos! Entre los odontocetos, el cachalote es el más grande, llegando a medir unos 20 metros. El cetáceo más pequeño es la vaquita marina, que mide alrededor de 140 centímetros.
| Especie | Longitud (m) | Peso (kg) | Notas |
|---|---|---|---|
| Vaquita marina | 1,2-1,5 | 30-55 | El cetáceo más pequeño. En peligro crítico de extinción. |
| Delfín común | 2,4 | 70-110 | |
| Delfín mular | 3,7 | 150-650 | |
| Narval | 5 | 800-1600 | |
| Ballena franca pigmea | 6,5 | 3000-3500 | El misticeto más pequeño. |
| Orca | 9,7 | 2600-9000 | El delfínido más grande. |
| Yubarta | 13,7-15,2 | 25 000-40 000 | |
| Cachalote | 14,9-20 | 13 000-14 000 | El odontoceto más grande. |
| Ballena azul | 30 | 110 000 | El animal más grande de todos. |
Piel y aletas
La piel de los cetáceos es muy gruesa, especialmente la capa de grasa que tienen debajo. Esta grasa les ayuda a no perder calor en el agua fría y también les sirve como reserva de energía.
Casi todos los cetáceos tienen una aleta dorsal en la espalda, que les da estabilidad al nadar. Su forma y tamaño varían mucho entre especies, lo que ayuda a identificarlas. La aleta caudal (la cola) es horizontal y se mueve de arriba abajo para impulsarlos. Las aletas pectorales, que son sus patas delanteras modificadas, les ayudan a girar y mantener el equilibrio.
¿Cómo funcionan los cetáceos?
Sentidos especiales
Los cetáceos tienen sentidos muy adaptados a la vida acuática:
- Vista: Sus ojos son planos y les permiten ver bien tanto bajo el agua como en el aire.
- Olfato: Está muy reducido en las ballenas con barbas y casi ausente en los delfines y cachalotes.
- Gusto: Tienen papilas gustativas en la lengua y pueden distinguir sabores. Los delfines mulares son muy sensibles al sabor salado, lo que podría ayudarles a orientarse.
- Tacto: Tienen muchos receptores táctiles en todo el cuerpo, especialmente en la cabeza y las aletas.
- Oído: Es su sentido más desarrollado. Pueden saber de dónde viene un sonido bajo el agua, algo que los mamíferos terrestres no pueden hacer. Esto es posible porque los huesos de su oído interno están separados del cráneo.
Respiración bajo el agua

Los cetáceos, como mamíferos, respiran aire con pulmones. Por eso, deben subir a la superficie para tomar O2 y expulsar CO2.
Sus narinas se han movido a la parte superior de la cabeza, formando el espiráculo. Esto les permite respirar casi completamente sumergidos. Los misticetos tienen dos orificios en el espiráculo, mientras que los odontocetos tienen solo uno. Ellos deciden cuándo respirar, a diferencia de nosotros.
Cuando exhalan, el aire caliente se condensa al contacto con el exterior, formando un chorro visible. La forma y altura de este chorro pueden ayudar a identificar la especie a distancia.
Los cetáceos pueden aguantar la respiración mucho más tiempo que otros mamíferos. Algunas especies, como el cachalote, pueden permanecer bajo el agua más de dos horas con una sola bocanada de aire. Esto se debe a que sus pulmones se colapsan a grandes profundidades, evitando problemas como la síndrome de descompresión que afecta a los buzos.
| Especie | Tiempo (min.) | Profundidad (metros) |
|---|---|---|
| Delfín del Pacífico de lados blancos | 5 | 210 |
| Delfín mular | 10 | 535 |
| Orca | 15 | 250 |
| Narval | 20 | 1000 |
| Yubarta | 20 | 150 |
| Ballena gris | 25 | 170 |
| Rorcual | 30 | 500 |
| Ballena azul | 50 | 100 |
| Ballena de Groenlandia | 80 | 300 |
| Ballenas pico de botella | 120 | 1.000 |
| Cachalote | 140 | 3.000 |
Corazón y sangre
El aparato circulatorio de los cetáceos es similar al de otros mamíferos. Su corazón tiene cuatro cámaras. Cuando se sumergen, su ritmo cardíaco disminuye (bradicardia), lo que ayuda a que la sangre llegue bien a órganos vitales como el cerebro y los riñones.
Sus músculos tienen mucha mioglobina, una proteína que almacena oxígeno, lo que les permite tener energía durante las inmersiones largas. Además, su sangre tiene una alta concentración de hemoglobina, que transporta eficientemente el oxígeno.
Esqueleto ligero

Los huesos de los cetáceos son ligeros y esponjosos, lo que les ayuda a flotar. Contienen mucha grasa. Su columna vertebral es muy flexible en la cola y la cabeza, permitiendo los movimientos que usan para nadar.
El cráneo de los cetáceos es especial. En los odontocetos, los huesos de la parte derecha del cráneo son más largos que los de la izquierda, lo que se relaciona con su capacidad de producir sonidos para la ecolocalización.
Sistema digestivo
El tracto digestivo de los cetáceos es muy largo. Su estómago tiene varias cámaras, como el de los rumiantes, lo que les ayuda a digerir sus alimentos.
Dientes y barbas: ¿Cómo se alimentan?

La principal diferencia entre odontocetos y misticetos es cómo comen:
- Odontocetos (con dientes): Sus dientes varían en forma y número. Algunos, como los cachalotes, solo tienen dientes en la mandíbula inferior. Los machos de narval son famosos por su largo colmillo en espiral, que usan en encuentros con otros machos.
- Misticetos (con barbas): Tienen barbas de queratina que cuelgan de su mandíbula superior. Estas barbas filtran el agua para atrapar pequeños peces y organismos. Las barbas más largas son las de las ballenas francas, que pueden medir hasta tres metros.
¿Cómo se reproducen los cetáceos?
Las crías de cetáceos se desarrollan dentro de la madre. Los ovarios de las hembras y los testículos de los machos están dentro del cuerpo para mantener la forma hidrodinámica.
En los odontocetos, el tiempo de desarrollo de la cría dentro de la madre dura entre 7 y 17 meses, dependiendo del tamaño del animal. En los misticetos, a pesar de ser más grandes, este periodo es más corto, entre 10 y 13 meses. Esto se debe a sus largas migraciones anuales.
Las crías nacen en el agua y son amamantadas por sus madres. Las madres y sus crías suelen formar fuertes lazos.
¿Cómo se mantienen calientes?
Los cetáceos son animales de sangre caliente y necesitan mantener su temperatura corporal constante. El agua enfría mucho más rápido que el aire, por lo que han desarrollado adaptaciones:
- Una gruesa capa de grasa bajo la piel actúa como un aislante térmico muy eficiente.
- En sus aletas y cola, tienen una red de vasos sanguíneos llamada rete mirabile que funciona como un intercambiador de calor, ayudándoles a regular su temperatura.
¿Cómo obtienen agua?
La mayoría de los cetáceos no beben agua de mar. Obtienen el agua que necesitan de los alimentos que comen o la producen a través de procesos químicos en su cuerpo al descomponer grasas y proteínas. Sus riñones están adaptados para manejar la sal del agua de mar.
¿Cuánto tiempo viven los cetáceos?
La mayoría de los cetáceos viven entre 20 y 30 años. Sin embargo, algunas especies viven mucho más. Se ha registrado que un rorcual común vivió 116 años, y algunas ballenas de Groenlandia pueden superar los 100 años, ¡llegando incluso a los 216 años en un caso!
¿Dónde viven los cetáceos?
Los cetáceos se encuentran en todos los mares y océanos del mundo. Algunas especies, como la orca, viven en casi todas las aguas. Otras, como la vaquita marina, viven en zonas muy específicas, como el golfo de California.
También hay especies que viven en agua dulce, en lagos y ríos de Norteamérica, Sudamérica y Asia. Algunos cetáceos prefieren las costas, mientras que otros viven en mar abierto.
¿Cómo se comportan los cetáceos?
Viajes y migraciones

Muchos misticetos realizan largas migraciones entre sus zonas de alimentación y sus zonas de reproducción. Por ejemplo, las yubartas se alimentan en las frías aguas polares en verano y viajan hacia el ecuador en invierno para reproducirse y tener a sus crías en aguas más cálidas y seguras.
Los odontocetos también migran, aunque las especies más pequeñas hacen viajes más cortos en busca de alimento. Se cree que los cetáceos usan el campo magnético de la Tierra, la posición del Sol o las corrientes marinas para orientarse en sus viajes.
¿Qué comen los cetáceos?
Todos los cetáceos son depredadores y están en la cima de la cadena trófica. Tienen pocos enemigos naturales, siendo el ser humano el más peligroso.
- Misticetos: Comen pequeños organismos como el plancton y peces pequeños, filtrándolos con sus barbas. Las yubartas a veces cazan en grupo, creando "redes de burbujas" para atrapar a los peces.
- Odontocetos: Comen presas más grandes, como cefalópodos (calamares) y peces. Las orcas son únicas porque también cazan otros mamíferos marinos.
Ecolocalización: el sonar de los odontocetos

Los odontocetos usan la ecolocalización para encontrar sus presas. Producen sonidos de alta frecuencia, llamados "clics", que viajan por el agua. Cuando estos sonidos chocan con algo, rebotan y el eco regresa a la mandíbula del animal, que lo transmite al oído. Así, pueden "ver" lo que hay a su alrededor, incluso en la oscuridad o en aguas turbias.
Algunos odontocetos también usan sonidos muy fuertes, llamados "bangs", para desorientar a sus presas. Los cachalotes y los zifios pueden sumergirse a grandes profundidades para cazar calamares, incluso calamares gigantes. Las orcas cazan en grupo, usando tácticas coordinadas para atrapar a sus presas.
Vida en grupo

Muchos cetáceos viven en grupos. Esto les ayuda a defenderse, cazar de forma más eficiente y para la reproducción.
- Misticetos: Generalmente no forman grupos muy grandes, excepto las yubartas, que se unen para cazar.
- Odontocetos: Tienen comportamientos sociales más complejos. Pueden formar grupos de miles de individuos, a veces incluso con diferentes especies. Las orcas, por ejemplo, viven en grupos liderados por la hembra más vieja, y los miembros se comunican con un "dialecto" propio del grupo.
¿Cómo se comunican?
Los cetáceos se comunican principalmente a través de sonidos, pero también usan el lenguaje corporal y el tacto.
Sonidos y cantos

Los odontocetos emiten silbidos y ladridos, además de los clics de ecolocalización. Algunos delfines tienen un "silbido firma" único para cada individuo. Las orcas tienen silbidos exclusivos de su grupo, formando un "dialecto" que aprenden las nuevas generaciones.
Los misticetos producen sonidos de baja frecuencia que pueden viajar muy lejos. Las yubartas son famosas por sus "cantos", que duran varios minutos y se repiten por horas. Estos cantos son complejos y se cree que los machos los usan para atraer a las hembras y comunicar su posición.
Comunicación táctil y corporal
Los cetáceos tienen muchas terminaciones nerviosas en su piel, especialmente en las aletas, la cabeza y otras zonas. Se frotan y acarician unos a otros con sus aletas pectorales, lo que podría fortalecer los lazos sociales en el grupo.
También se comunican con movimientos corporales, como golpear la cola o las aletas contra el agua, o saltar fuera de ella. Estos comportamientos pueden tener diferentes significados, como mostrar hostilidad, jugar o incluso quitarse parásitos.
¿Cómo se relacionan con los humanos?

Desde la antigüedad, los delfines han sido vistos como animales especiales. Los griegos los asociaban con el dios Apolo y hay historias de delfines que ayudaron a pescadores o salvaron a personas de ahogarse. Hoy en día, en algunos lugares, como en Laguna, Brasil, los delfines colaboran con los pescadores para atrapar peces, beneficiándose ambos.
Los cetáceos también han aparecido en muchas obras de arte y entretenimiento. La famosa novela Moby Dick trata sobre la caza de una ballena blanca. Películas como Liberen a Willy han mostrado la amistad entre humanos y orcas.

En los últimos años, el avistamiento de cetáceos se ha vuelto muy popular. Es una actividad turística que permite a las personas observar a estos animales en su ambiente natural. Esto es importante para el turismo, la investigación y la protección de los cetáceos.
¿Cuál es el estado de conservación de los cetáceos?
Muchos cetáceos están en peligro. Según la Lista Roja de la UICN, catorce especies están seriamente amenazadas. Dos de ellas, el delfín de río chino y la vaquita marina, están en peligro crítico de extinción. El delfín de río chino se considera casi extinto.
Las principales amenazas para los cetáceos son:
- Caza: La caza de ballenas ha existido por miles de años. Antes se cazaban por su grasa y barbas, ahora por su carne en algunos países. Esto ha reducido drásticamente sus poblaciones.
- Capturas accidentales: Muchos cetáceos mueren al quedar atrapados en redes de pesca.
- Contaminación: Sustancias químicas y metales pesados en el agua pueden afectar su salud y reproducción. Los derrames de petróleo también son peligrosos.
- Ruidos submarinos: El ruido de barcos, sonares militares y otras actividades humanas bajo el agua puede interferir con su comunicación y ecolocalización, causando daños en su oído.
- Enfermedades: Son sensibles a ciertos virus y toxinas.
- Colisiones con barcos: Los cetáceos grandes pueden chocar con embarcaciones, especialmente si están descansando en la superficie.
- Pérdida de hábitat: La destrucción de sus hogares naturales.
Varadas: ¿Por qué los cetáceos llegan a la orilla?

A veces, los cetáceos, especialmente los calderones negros y los cachalotes, quedan varados en la orilla y no pueden volver al mar. Esto puede causarles la muerte por deshidratación o asfixia. Las varadas pueden ser de un solo animal o de un grupo entero.
Las causas de las varadas no están del todo claras, pero se cree que son una combinación de factores naturales (cambios en corrientes, tormentas), biológicos (enfermedades, depredadores) y de comportamiento. En especies muy sociales, si un animal se vara, otros miembros del grupo pueden seguirlo y también quedar atrapados.
Se ha investigado la relación entre las varadas y la actividad solar, así como con el uso de sonares militares. Se ha encontrado una conexión entre las varadas masivas de zifios y las operaciones militares con sonar en la zona.
¿Qué es la cetología?

La cetología es la rama de la biología marina que se dedica a estudiar a los cetáceos. Gran parte de lo que sabemos sobre ellos proviene del estudio de animales varados.
Desde la antigüedad, pensadores como Aristóteles ya estudiaban a los cetáceos. Aristóteles los diferenció de los peces, reconociendo que eran mamíferos que respiraban aire y amamantaban a sus crías. También notó la diferencia entre los cetáceos con barbas y los que tenían dientes.
A lo largo de la historia, el conocimiento sobre los cetáceos ha crecido. Hoy en día, los científicos usan tecnología como radiotransmisores satelitales, hidrófonos para escuchar sus sonidos y cámaras para identificarlos, ya que pasan la mayor parte de su tiempo bajo el agua.
Galería de imágenes
-

Diferentes tipos de cetáceos.
-

Pakicetus, un antiguo pariente de los cetáceos.
-
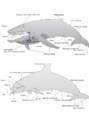
Descripción de las características de un misticeto (arriba) y un odontoceto (abajo).
-

Respiración de un rorcual azul.
-

Esqueleto de ballena azul.
-
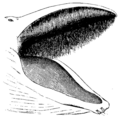
Ilustración de la boca de una ballena con barbas.
-

Aleta caudal del cachalote Mediterráneo Amanita. Las aletas caudales y dorsales permiten seguir a largo plazo a los cetáceos. Amanita lleva 20 años visitando el Estrecho.
-

Un grupo de yubartas se alimentan usando una red de burbujas.
-

Una ballena franca glaciar con su cría.
-
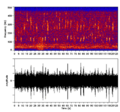
Espectrograma del canto de una yubarta.
-

Moneda tarentina del 500-480 a. C. que presenta a Taras, fundador de la ciudad, cabalgando un delfín.
-

Turistas observando cetáceos. Esta actividad, llamada avistamiento de cetáceos, es cada vez más popular.
-

Voluntarios ayudando a calderones negros varados en Nueva Zelanda.
-

1- Ballena de Groenlandia; 2- Orca; 3- Ballena franca; 4- Cachalote; 5- Narval; 6- Ballena azul; 7- Rorcual; 8- Beluga.
-

Aleta dorsal de una orca (Orcinus orca).
-

Aleta caudal de un cachalote.
-

Aleta pectoral de una yubarta.
-

Cráneo de orca.
-
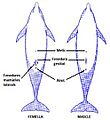
Disposición de las hendiduras en el delfín mular.
-

Madre e hijo de orca en el Estrecho.
-

Detalle de marca satélite en cachalote.
-

Grupo de delfines mulares indo pacíficos en el mar Rojo.
-

Una orca hace spyhopping en un agujero en el hielo.
-

Delfines de flancos blancos del Atlántico capturados y muertos en una batida de caza en las islas Feroe.
-

Una marsopa atrapada en una red de pescar.









.jpg)

Véase también
 En inglés: Cetacea Facts for Kids
En inglés: Cetacea Facts for Kids
