
Carnotaurus sastrei para niños
Datos para niños Carnotaurus sastrei |
||
|---|---|---|
| Rango temporal: 72 Ma - 69,9 Ma Cretácico superior | ||
.png) Reconstrucción en vida.
|
||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Ceratosauria | |
| Superfamilia: | Abelisauroidea | |
| Familia: | Abelisauridae | |
| Subfamilia: | Carnotaurinae | |
| Tribu: | Carnotaurini | |
| Género: | Carnotaurus Bonaparte, 1985 |
|
| Especie: | C. sastrei Bonaparte, 1985 |
|
El Carnotaurus sastrei (que significa "toro carnívoro de Anselmo Sastre" en latín), o carnotauro en español, es un dinosaurio terópodo extinto. Es la única especie conocida de su género, Carnotaurus, que pertenece a la familia Abelisauridae. Vivió en el Cretácico superior, hace unos 72 a 69.9 millones de años, en lo que hoy es Sudamérica.
Lo más llamativo del carnotauro son sus dos pequeños cuernos sobre los ojos. Fue el primer dinosaurio carnívoro conocido con cuernos bien definidos. También se encontraron impresiones de su piel, que mostraban pequeños escudos óseos llamados osteodermos. El carnotauro era un miembro avanzado de los abelisáuridos. Este grupo de grandes terópodos fue el principal depredador en las tierras que formaban Gondwana al final del Cretácico.
Junto con el género Abelisaurus, el carnotauro ayudó a los científicos a entender una nueva familia de depredadores de la era Mesozoica, los abelisáuridos. Esto cambió la forma en que se clasificaban otros dinosaurios y dio una nueva visión de la vida animal en Gondwana al final de la era de los dinosaurios. El nombre "sastrei" se le dio en honor a Anselmo Sastre, dueño de la estancia en Chubut, Argentina, donde se encontró el primer y único fósil.
Los científicos aún no están seguros de qué comía el carnotauro. Algunos estudios de su cráneo sugieren que podía cazar presas muy grandes, como los saurópodos. Otros estudios indican que se alimentaba principalmente de animales más pequeños. Lo que sí se sabe es que el carnotauro era muy rápido. Posiblemente fue uno de los terópodos grandes más veloces, alcanzando velocidades de hasta 56 kilómetros por hora. Podía medir hasta 3.5 metros de alto y 7.6 metros de largo.
Contenido
¿Cómo era el Carnotauro?

El carnotauro era un terópodo de tamaño mediano a grande, pero de cuerpo ligero. Medía entre ocho y nueve metros de largo y unos 3.5 metros de alto. Se estima que pesaba entre 1350 y 2100 kg. Esto lo convierte en uno de los abelisáuridos más grandes conocidos. Solo Pycnonemosaurus y quizás Abelisaurus pudieron ser más grandes, pero sus restos son incompletos.
Su cabeza era muy pequeña en comparación con otros depredadores de su tamaño, midiendo unos sesenta centímetros de largo. Las características más distintivas del carnotauro son sus dos cuernos gruesos sobre los ojos. También tenía unas patas delanteras muy pequeñas y atrofiadas, con cuatro dedos. Estudios muestran que el carnotauro era un terópodo muy especializado. Esto se ve en su cráneo, vértebras y patas delanteras. Sin embargo, su pelvis y patas traseras eran más simples, parecidas a las del Ceratosaurus. Los huesos de su muslo eran largos y delgados.
La cabeza del Carnotauro

El carnotauro tenía un cráneo fuerte pero pequeño, con espacios vacíos para reducir el peso. Tenía un pecho grueso y una cola larga y robusta que le ayudaba a mantener el equilibrio. Sus ojos eran pequeños y estaban protegidos. Miraban hacia adelante, lo cual es raro en un dinosaurio. Esto podría significar que tenía visión binocular y podía calcular bien las distancias. Su hocico grande sugiere que tenía un sentido del olfato muy desarrollado. Sus mandíbulas podían abrirse mucho para morder.
El cráneo medía 59.6 cm de largo y era más corto y profundo que el de otros dinosaurios carnívoros grandes. El hocico era ancho y las mandíbulas se curvaban hacia arriba. Como en otros abelisáuridos, los huesos de su cara tenían muchos pequeños agujeros y espinas. En vida, una piel gruesa y posiblemente con queratina cubría estos huesos. Un par de cuernos prominentes sobresalían por encima de sus ojos. Estos cuernos, hechos de los huesos frontales, eran gruesos y planos en la parte superior, midiendo quince cm de largo. Probablemente tenían una cubierta de queratina que los hacía aún más grandes.
Los ojos, pequeños, estaban en la parte superior de las cuencas oculares, que tenían forma de cerradura. La parte superior de la cuenca estaba ligeramente girada hacia adelante, lo que permitía cierta visión binocular. La mandíbula inferior del carnotauro tenía una conexión débil entre sus huesos. Esto formaba una gran abertura en la mandíbula. Los dientes eran largos y delgados, a diferencia de los dientes cortos de otros abelisáuridos. Tenía cuatro dientes en la parte delantera de la mandíbula superior y doce en los lados. La mandíbula inferior tenía quince dientes a cada lado.

El cráneo del carnotauro tenía seis aberturas principales a cada lado, como en otros dinosaurios. La abertura más frontal, la fosa nasal ósea, era rectangular y miraba hacia los lados y hacia adelante. Entre la fosa nasal y la cuenca del ojo estaba la fenestra antorbitaria. En el carnotauro, esta abertura era más alta que larga. El ojo estaba en la parte superior de la cuenca ocular, que tenía forma de cerradura. Esta parte superior era pequeña y circular.
La mandíbula inferior del carnotauro era delgada y no muy fuerte. Sin embargo, estudios más recientes sugieren que las conexiones entre los huesos de la mandíbula eran flexibles, lo que la hacía adaptable pero no necesariamente débil. Se encontraron huesos hioides osificados en la mandíbula inferior. Estos huesos delgados sostienen los músculos de la lengua y otros músculos. Rara vez se encuentran en dinosaurios porque no están conectados a otros huesos y se pierden fácilmente. El carnotauro es el único terópodo no aviar conocido con un hueso basihyal.
La columna vertebral del Carnotauro

La columna vertebral del carnotauro tenía diez vértebras cervicales en el cuello, doce dorsales en la espalda, seis sacras fusionadas y un número desconocido de vértebras en la cola. Su cuello era casi recto, no con la curva en "S" que se ve en otros terópodos. También era inusualmente ancho, especialmente en la base. La parte superior de la columna del cuello tenía dos filas de huesos agrandados llamados epipófisis, que hacían que la parte superior fuera plana. Estos huesos eran los puntos más altos de la columna y probablemente servían para unir músculos del cuello muy fuertes. Una doble fila similar también estaba presente en la cola, formada por costillas caudales modificadas en forma de "V", lo que resultaba en una parte superior plana de la cola.
La piel del Carnotauro

Solo se ha encontrado un esqueleto de carnotauro, casi completo. Incluye impresiones de la piel a lo largo de casi todo el lado derecho. Estas impresiones no muestran evidencia de plumas u otras estructuras filamentosas. La piel tenía pequeños osteodermos, que eran escudos óseos. Estos escudos tenían forma de conos de unos cinco centímetros de diámetro. Aumentaban de tamaño a medida que se acercaban a la línea central de la espalda. Esto le daba un aspecto similar al de un cocodrilo actual, sugiriendo que el carnotauro tenía una piel áspera y gruesa.
El carnotauro fue el primer dinosaurio terópodo descubierto con impresiones completas de piel fosilizada. Estas impresiones se encontraron debajo del lado derecho del esqueleto y provienen de diferentes partes del cuerpo. La mayor parte de la piel corresponde a la parte delantera de la cola. La piel estaba formada por un mosaico de escamas poligonales que no se superponían y medían unos cinco mm de diámetro. Este mosaico estaba dividido por finas ranuras paralelas. El patrón de las escamas era similar en todo el cuerpo, excepto en la cabeza, que parecía tener un patrón diferente con escamas irregulares.

De manera única para los terópodos, el carnotauro tenía osteodermos (escudos óseos) a lo largo de los lados del cuello, la espalda y la cola en filas irregulares. Cada uno de estos escudos tenía una cresta baja y medía entre cuatro y cinco cm de diámetro. Estaban espaciados entre ocho y diez cm y eran más grandes hacia la parte superior del animal. Se ha sugerido que estas estructuras podrían haber protegido al animal durante peleas con otros carnotauros o terópodos.
Los brazos del Carnotauro

Sus patas delanteras eran más cortas que las del Tyrannosaurus. Tenían cuatro dedos en cada mano, lo cual se considera una característica primitiva en los dinosaurios. No tenían huesos del carpo en la mano, por lo que los metacarpianos se unían directamente al antebrazo. La mano tenía los cuatro dedos básicos, aunque solo los dos del medio terminaban en huesos de los dedos. El cuarto dedo era solo un metacarpiano en forma de espolón. Se cree que este último se usaba para sujetar a las presas. Los dedos estaban fusionados e inmóviles, y podrían no haber tenido garras.
El carnotauro se diferenciaba de otros abelisáuridos por tener patas delanteras proporcionalmente más cortas y robustas. También tenía el cuarto metacarpo como el hueso más largo de la mano. La proporción de sus patas delanteras con respecto a las traseras era del 14%, mucho menor que en otros terópodos. El húmero medía 285 milímetros, el radio 73 mm y el metacarpo 37 mm. Esto muestra que el carnotauro tenía la reducción más marcada de las patas delanteras entre los terópodos.

Algunos investigadores han sugerido que los huesos del brazo del carnotauro podían girar 180 grados, dejando las palmas de las manos hacia afuera. Un estudio de 2009 sugiere que los brazos de los abelisáuridos eran vestigiales (sin función aparente). Esto se debe a que las fibras nerviosas que transmiten los estímulos nerviosos estaban muy reducidas, similar a lo que se ve hoy en aves como los emúes y los kiwis, que también tienen extremidades delanteras vestigiales.
Descubrimiento e investigación del Carnotauro

El carnotauro fue descubierto en la Patagonia de Argentina, en la Provincia del Chubut. El esqueleto, conocido como MACN-CH 894, fue hallado en 1984 por una expedición liderada por el paleontólogo argentino José Fernando Bonaparte. Sus restos se encontraron en rocas de la Formación La Colonia. Este hallazgo fue muy importante porque dio a los científicos pistas sobre cómo era la piel de los dinosaurios. En esta expedición también se encontró el saurópodo con espinas Amargasaurus.
El esqueleto está bien conservado y articulado, lo que significa que los huesos aún estaban conectados. Solo le faltaban los dos tercios posteriores de la cola, la parte inferior de la pierna y los pies, que fueron dañados por el clima. El cráneo y el hocico se aplastaron durante la fosilización. El esqueleto pertenecía a un carnotauro adulto. Se encontró acostado sobre su lado derecho, con el cuello doblado hacia atrás, una postura común en los fósiles. Lo inusual es que se conservaron extensas impresiones de su piel. Debido a la importancia de estas impresiones, se realizó una segunda expedición para buscar más restos de piel.
El fósil fue encontrado en la estancia "Pocho Sastre", cerca de Bajada Moreno, en Chubut, Argentina. Los sedimentos corresponden a la parte inferior de la Formación La Colonia, del Cretácico superior, con una antigüedad de unos 72 a 68 millones de años. El fósil original de Carnotaurus sastrei se guarda en el Museo Argentino de Ciencias Naturales Bernardino Rivadavia, donde también se puede ver una réplica de su esqueleto. La preparación del fósil fue difícil porque estaba incrustado en una roca muy dura llamada hematites.
En 1985, Bonaparte publicó una nota presentando a Carnotaurus sastrei como un nuevo género y especie. Describió brevemente el cráneo y la mandíbula inferior. El nombre del género, Carnotaurus, viene del latín "carnis" (carne) y "taurus" (toro), refiriéndose a sus cuernos. El nombre de la especie, sastrei, honra a Ángel Sastre, el dueño del rancho. Una descripción completa del esqueleto se hizo en 1990. Después de Abelisaurus, el carnotauro fue el segundo miembro de la familia Abelisauridae en ser descubierto. Durante años, fue el abelisáurido más completo y el terópodo mejor conocido del hemisferio sur. No fue hasta el siglo XXI que se describieron otros abelisáuridos bien conservados, como Aucasaurus, Majungasaurus y Skorpiovenator. Esto permitió a los científicos reevaluar algunos aspectos de la anatomía del carnotauro.
¿Cretácico inferior o superior?

Al principio, se pensó que el carnotauro era del Cretácico medio, hace cien millones de años. Sin embargo, los primeros estudios ya decían que las fechas de las rocas donde se encontró no eran muy precisas. En 2002, M. C. Lamanna y sus colegas publicaron una fecha definitiva. Esta fecha sitúa al carnotauro a finales del Cretácico superior, en la Formación La Colonia. Esto significa que vivió al mismo tiempo que otros abelisáuridos con cuernos de la India y Madagascar.
Clasificación del Carnotauro
El carnotauro es uno de los géneros mejor conocidos dentro de los Abelisauridae. Esta familia de grandes terópodos se encontraba en el antiguo supercontinente Gondwana y en algunas partes de lo que hoy es Europa. Los abelisáuridos eran los depredadores dominantes en el Cretácico Tardío en estas áreas. Reemplazaron a otros grandes carnívoros y ocuparon el lugar que los tiranosáuridos tenían en los continentes del norte.
Aunque el cráneo del carnotauro es muy diferente al de Abelisaurus comahuensis, ambos tienen características que los clasifican como Abelisauridae. Estas incluyen una gran abertura infratemporal, un hueso cuadrado alargado y otras características específicas del cráneo. Por estas razones, José Fernando Bonaparte, Fernando Novas y Rodolfo Coria propusieron que Abelisaurus y Carnotaurus pertenecen a una nueva familia llamada Abelisauridae.
Se han observado grandes diferencias entre los terópodos de Laurasia (continentes del norte) y los de Gondwana (continentes del sur). Estas diferencias se ven en el cráneo, la columna vertebral y la notable reducción de las patas delanteras. Se cree que estas diferencias anatómicas se deben a la separación geográfica de los supercontinentes al final del Cretácico. El carnotauro da nombre a dos subgrupos dentro de Abelisauridae: la subfamilia Carnotaurinae y la tribu Carnotaurini.
Taxonomía del Carnotauro
Carnotaurus sastrei es la única especie conocida de su género. Fue descubierta en 1985 por el paleontólogo José F. Bonaparte en la provincia argentina de Chubut. Sus restos se encontraron en rocas de la Formación La Colonia. Pertenece a la familia Abelisauridae. Aunque las relaciones dentro de los Abelisauridae se debaten, el carnotauro se considera uno de los miembros más avanzados de la familia en los análisis cladisticos. Su pariente más cercano podría haber sido Aucasaurus o Majungasaurus.
En 2008, Canale y sus colegas crearon el clado Brachyrostra. Este grupo incluye al carnotauro y otros carnotaurinos de Sudamérica. En 2015, Filippi y sus colegas crearon un nuevo clado, Furileusauria, que incluye a los Brachyrostra más avanzados. Esto coloca al carnotauro solo en Carnotaurinae, mientras que Aucasaurus y Abelisaurus están en la subfamilia Abelisaurinae.
Paleobiología del Carnotauro

El carnotauro es el único carnívoro bípedo conocido con un par de cuernos frontales robustos. Se ha sugerido que el carnotauro usaba su cráneo para impactos de alta velocidad al cazar. Otra idea es que los cuernos se usaban en combates con otros carnotauros por territorio, reproducción o liderazgo. Sin embargo, un estudio más reciente mostró que un carnotauro no podría haber sobrevivido a un choque frontal con sus cuernos.
Se ha estudiado que el carnotauro era muy rápido, alcanzando unos cincuenta km/h. Se le considera el terópodo más veloz que haya existido. La fuerza de su cráneo se complementaba con músculos del cuello poderosos y una columna vertebral rígida. Al igual que el Allosaurus, el carnotauro tenía adaptaciones para cazar presas más grandes que él. Tenía articulaciones en la mandíbula que le permitían abrir la boca de forma excepcional. Sus músculos del cuello le daban más fuerza para mover la cabeza.
La mordida y el cráneo del Carnotauro
Los estudios de la mandíbula del carnotauro sugieren que podía morder rápidamente, pero no con mucha fuerza. Estos movimientos rápidos son importantes para atrapar presas pequeñas, como se ve en los cocodrilos actuales. Los investigadores también notaron que el cráneo era muy flexible, especialmente la mandíbula inferior, similar a las serpientes modernas. Esta elasticidad le habría permitido al carnotauro tragar presas pequeñas enteras.
La parte delantera de la mandíbula inferior podía moverse hacia arriba y hacia abajo. Cuando se movía hacia abajo, los dientes se proyectaban hacia adelante para atrapar presas. Cuando se movía hacia arriba, los dientes se proyectaban hacia atrás, lo que impedía que la presa escapara. Sin embargo, los mismos investigadores también encontraron que el cráneo podía soportar las fuerzas al tirar de presas grandes. Esto sugiere que el carnotauro no solo comía presas pequeñas, sino que también podía cazar dinosaurios grandes.

A pesar de su aspecto ligero, el cráneo del carnotauro era muy resistente. La fuerza máxima que podía resistir era mucho mayor que la fuerza de su mordida. Esto le daba un factor de seguridad muy alto, lo que sugiere que su cráneo podría haber sido "sobrediseñado". Los cuernos del carnotauro eran anchos y planos en la parte superior, lo que proporcionaba una base de contacto amplia. La presencia de cuernos y la fuerte unión de los huesos frontales y parietales sugieren que esta parte del cráneo estaba diseñada para resistir grandes fuerzas de compresión.
La fuerza máxima de mordida estimada en los dientes traseros era de 3341 N. Esto es relativamente débil en comparación con otras especies y no habría sido suficiente para triturar huesos de presas medianas o grandes. El gran rango de movimientos y el diseño de sus dientes, junto con la fuerza de los músculos de su cráneo y cuello, le habrían permitido al carnotauro cortar y desgarrar grandes trozos de alimento para tragarlos sin masticar.
François Therrien y sus colegas cuestionaron la idea de que el carnotauro solo cazaba presas pequeñas. Encontraron que la fuerza de mordida del carnotauro era el doble que la del cocodrilo americano, que tiene una de las mordidas más fuertes entre los tetrápodos vivos. También notaron similitudes con los modernos dragones de Komodo. La resistencia a la flexión de la mandíbula inferior disminuía hacia la punta. Esto indica que las mordidas no eran adecuadas para la precisión necesaria al cazar presas pequeñas. Sin embargo, podían usarse para herir y debilitar a una presa grande. Por lo tanto, el carnotauro probablemente cazaba animales grandes, quizás emboscándolos.
Robert Bakker sugirió que el carnotauro se alimentaba principalmente de presas muy grandes, especialmente saurópodos. Varias adaptaciones de su cráneo (hocico corto, dientes pequeños y parte trasera del cráneo fuerte) también evolucionaron en el Allosaurus. Estas características sugieren que la mandíbula superior se usaba como un hacha para infligir heridas a grandes saurópodos, que se debilitarían con ataques repetidos.
Función de los cuernos del Carnotauro
Hasta ahora, el carnotauro es el único carnívoro conocido con un par de cuernos en el hueso frontal. El uso de estos cuernos no está del todo claro. Hay varias ideas: podrían haberse usado en peleas con otros carnotauros, para exhibirse o para matar presas.

Gregory S. Paul propuso que los cuernos se usaban en batallas de empujones. Sugirió que la cuenca del ojo se redujo para minimizar el daño a los ojos durante la lucha. Gerardo Mazzetta y sus colegas sugirieron que el carnotauro usaba los cuernos de manera similar a los carneros actuales. Calcularon que los músculos del cuello eran lo suficientemente fuertes para absorber la fuerza de dos individuos chocando de frente a una velocidad de 5.7 m/s cada uno. Fernando Novas interpretó varias características del esqueleto como adaptaciones para dar golpes con la cabeza. Sugirió que el cráneo pequeño permitía movimientos de cabeza más rápidos, y el cuello musculoso permitía golpes fuertes.
Otros estudios sugieren que el carnotauro no podía dar golpes rápidos con la cabeza. En cambio, podría haberse empujado lentamente con otros carnotauros usando los lados superiores de sus cráneos. Por lo tanto, los cuernos podrían haber sido un dispositivo para disipar las fuerzas de compresión y proteger el cerebro. Esto se apoya en los lados superiores aplanados de los cuernos, los huesos fuertemente fusionados de la parte superior del cráneo y la incapacidad del cráneo para soportar golpes rápidos en la cabeza.
Mazzetta y sus colegas también propusieron que los cuernos podrían haberse usado para herir o matar presas pequeñas. Aunque los núcleos óseos eran romos, podrían haber tenido una cubierta de queratina similar a la de los bóvidos modernos. Sin embargo, esta teoría es menos creíble, ya que sería el único caso conocido de cuernos usados como armas de caza en animales.
¿Cómo se movía el Carnotauro?
Mazzetta y sus colegas creen que el carnotauro era un corredor veloz. Argumentan que el hueso del muslo estaba adaptado para soportar grandes fuerzas al correr. La capacidad de la pierna de un animal para soportar estas fuerzas limita su velocidad máxima. Las adaptaciones del carnotauro eran superiores a las de un ser humano, aunque no tan buenas como las de un avestruz, ambos bípedos. Los científicos calculan que el carnotauro podía alcanzar una velocidad máxima de 48 a 56 kilómetros por hora.

En los dinosaurios, el músculo más importante para moverse se encuentra en la cola. Este músculo, llamado caudofemoralis, se une a una cresta prominente en el fémur (hueso del muslo) y tira del hueso del muslo hacia atrás al contraerse. Scott Persons y Philip J. Currie señalan que en las vértebras de la cola del carnotauro, las costillas caudales no sobresalían horizontalmente, sino en ángulo, formando una "V". Esto habría proporcionado espacio adicional para un músculo caudofemoralis más grande que en cualquier otro terópodo. Se calculó que la masa muscular era de entre 111 y 137 kilogramos por pierna. Por lo tanto, el carnotauro podría haber sido uno de los terópodos grandes más rápidos conocidos.
Mientras que el músculo caudofemoralis era grande, los músculos situados por encima de las costillas caudales habrían sido proporcionalmente más pequeños. Estos músculos eran responsables del movimiento y la estabilidad de la cola. Para mantener la estabilidad de la cola a pesar de la reducción de estos músculos, las costillas caudales tenían proyecciones que se entrelazaban con las vértebras y la pelvis. Esto aumentaba la rigidez de la cola. Como resultado, la capacidad de girar rápidamente habría disminuido, ya que la cadera y la cola tenían que moverse al mismo tiempo, a diferencia de otros terópodos.
Cerebro y sentidos del Carnotauro
Cerroni y Paulina-Carabajal, en 2019, usaron tomografías computarizadas para estudiar la cavidad que contenía el cerebro. El volumen de esta cavidad era de 168.8 cm³, aunque el cerebro solo habría llenado una parte de este espacio. Los bulbos olfativos, que controlaban el sentido del olfato, eran grandes. En cambio, los lóbulos ópticos, responsables de la vista, eran relativamente pequeños. Esto indica que el sentido del olfato del carnotauro podría haber sido mejor que su vista.
La parte frontal de los tractos olfatorios y los bulbos se curvaba hacia abajo, una característica que solo comparte Indosaurus. En otros abelisáuridos, estas estructuras estaban horizontales. Según Cerroni y Paulina-Carabajal, esta curvatura y el gran tamaño de los bulbos podrían indicar que el carnotauro dependía más de su olfato que otros abelisáuridos. El flóculo, una parte del cerebro relacionada con la estabilización de la mirada y la coordinación entre los ojos y el cuerpo, era grande en el carnotauro y otros abelisáuridos sudamericanos. Esto podría indicar que estos dinosaurios usaban con frecuencia movimientos rápidos de cabeza y cuerpo. La audición podría haber estado poco desarrollada en el carnotauro, como indica la lagena corta de su oído interno. Se estimó que su rango de audición era inferior a 3 kHz.
Paleoecología del Carnotauro

La Formación La Colonia se interpreta como depósitos de un ambiente de estuarios. Estos eran lugares con costas bajas donde el agua dulce del continente se mezclaba con las mareas marinas. Se han encontrado moluscos bivalvos similares a ostras. La mayoría de los vertebrados encontrados incluyen peces, tortugas, crocodilomorfos, plesiosauros, dinosaurios, serpientes y mamíferos.
Entre los dinosaurios, además del carnotauro, se encontraron fragmentos de otros terópodos, una vértebra de un titanosauriano, partes de un hadrosauroide y osteodermos de un anquilosauriano. En 2024, se nombró al segundo dinosaurio no aviar de la formación, el saltasauroide Titanomachya gimenezi. Entre las serpientes se encontraron de las familias Boidae y Madtsoidae. Las tortugas están representadas por al menos cinco tipos. También se encontraron restos de peces dipnoos, otros peces de agua dulce y anuros.
En la Patagonia, y viviendo al mismo tiempo que el carnotauro, habitaron otros dinosaurios. Entre ellos estaban los titanosaurianos Antarctosaurus, Puertasaurus, Neuquensaurus, Epachthosaurus, Pellegrinisaurus, Rocasaurus, Aeolosaurus, Bonatitan, Laplatasaurus y Saltasaurus. También los euiguanodóntidos Talenkauen y Gasparinisaura, el hadrosáurido Secernosaurus y el posible ceratopsiano Notoceratops. Todos estos eran herbívoros y posibles presas del carnotauro. También compartió el territorio con el gran megarraptórido Orkoraptor, los abelisáuridos Quilmesaurus y Aucasaurus, y probablemente con el gran dromeosáurido Austroraptor.
El Carnotauro en la cultura popular
Hasta mediados de los años 1990, el carnotauro no aparecía mucho en los medios. Una de sus primeras apariciones importantes fue en 1995, en la novela El mundo perdido, la secuela de la exitosa novela Parque Jurásico de Michael Crichton. En la novela, el carnotauro tenía la habilidad de cambiar de color como un camaleón o una sepia, lo que le permitía camuflarse. Esto es una idea del escritor y no se basa en pruebas científicas. Esta característica fue omitida en la versión cinematográfica de 1997.
El Carnotauro en el cine
Aparece en la película de Disney, Dinosaurio (2000). Allí, una pareja de carnotauros son los principales villanos, persiguiendo a una manada de dinosaurios herbívoros. En la película, el carnotauro aparece mucho más grande de lo que era en realidad, con un tamaño similar al de un Tiranosaurio. En verdad, el carnotauro tenía un tamaño parecido al del Iguanodonte.
Galería de imágenes
-

Montura del esqueleto de un carnotauro.
-

Cráneo de un carnotauro desde un punto de vista lateral.
-

Montura anacrónica de un carnotauro y un amargasaurio.
-
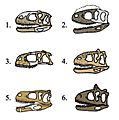
Comparativa de cráneos de abelisáuridos: 1)Indefinido 2)Rugops 3)Abelisaurus 4)Majungasaurus 5)Aucasaurus 6)Carnotauro.
-
Dibujos de dos carnotauros.





Véase también
 En inglés: Carnotaurus Facts for Kids
En inglés: Carnotaurus Facts for Kids
- El mundo perdido (novela)
- Dinosaurio (película)
- Abelisauridae
- Abelisaurus
- José Fernando Bonaparte
- Anexo:Géneros válidos de dinosaurios
