
Coral para niños
Datos para niños
Coral |
||
|---|---|---|
 Coral en forma de pilar (Dendrogyra cylindrus)
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Cnidaria | |
| Clase: | Anthozoa Ehrenberg, 1831 |
|
| Subclases y son animales vivientes órdenes existentes | ||
|
Subclase: Octocorallia |
||
Los corales marinos son animales coloniales, salvo excepciones, pertenecientes al filo Cnidaria, la mayoría a la clase Anthozoa aunque algunos son de la clase Hydrozoa (como Millepora). Los corales están formados por cientos o miles de individuos llamados zooides y pueden alcanzar grandes dimensiones. En aguas tropicales y subtropicales forman grandes arrecifes. El término «coral» no tiene significado taxonómico y bajo él se incluyen diferentes tipos de organismos.
Contenido
Características
Aunque los corales pueden atrapar plancton y pequeños peces ayudados por las células urticantes de sus tentáculos, la mayoría de los corales obtienen la mayor parte de sus nutrientes de las algas unicelulares fotosintéticas, denominadas zooxantelas, que viven dentro del tejido del coral y le dan color a este. Estos corales requieren de luz solar y crecen en agua clara y poco profunda, normalmente a profundidades menores de 60 metros. Los corales pueden ser los principales contribuyentes a la estructura física de los arrecifes de coral que se formaron en aguas tropicales y subtropicales, como la enorme Gran Barrera de Coral en Australia y el arrecife Mesoamericano en el mar Caribe. Otros corales, que no tienen una relación simbiótica con algas, pueden vivir en aguas mucho más profundas y en temperaturas mucha más bajas, como las especies del género Lophelia que pueden sobrevivir hasta una profundidad de 3000 metros.
El término "coral" no tiene ningún significado taxonómico y es poco preciso; suele usarse para designar a los antozoos en general, tanto a los que generan un esqueleto calcáreo duro, especialmente los que construyen colonias ramificadas, como las acroporas; pero también es común denominar coral a especies con colonias compactas ("coral cerebro" como Lobophyllia) e incluso con esqueleto córneo y flexible, como las gorgonias. Asimismo, se llaman corales blandos a las especies del orden Alcyonacea, que no generan esqueleto y utilizan el calcio en forma de espículas repartidas por su tejido carnoso, para proporcionarles mayor rigidez y consistencia.
Tanto en el mundo del buceo como en acuariofilia, los corales se dividen en blandos y duros, según tengan esqueleto o no. Y los duros, a su vez, se subdividen en duros de pólipo corto y duros de pólipo largo, "small polyp stony" (SPS) y "large polyp stony" (LPS), en inglés. Esta clasificación se refiere al tamaño del pólipo, que en los SPS como Montipora, suele ser de 1 o 2 mm y en los LPS, como Heliofungia actiniformis de hasta 20 cm.Pero esta división es poco científica y genera no pocas excepciones bajo un análisis riguroso de las diversas especies. La comunidad científica se refiere a micropólipos cuando los pólipos coralinos tienen entre 1 y 2 mm de diámetro, y alude a macropólipos para aquellos pólipos entre 10 y 40 mm de diámetro. No obstante, la gran mayoría de los corales de todos los arrecifes del mundo tienen pólipos con un diámetro entre 2 y 10 mm, justo entre esas categorías.
Clasificación de los corales


Los corales se dividen en dos subclases en función del número de tentáculos o líneas de simetría: Hexacorallia y Octocorallia, y una serie de órdenes en función de sus exoesqueletos, tipo de nematocisto y análisis genético mitocondrial. La tipificación común de los corales cruza los límites del suborden/clase.
Corales hermatípicos
Los corales hermatípicos, son corales pétreos que construyen arrecifes. Secretan carbonato de calcio para formar un esqueleto duro. Obtienen una parte de sus requerimientos energéticos de zooxantelas, (algas fotosintéticas simbióticas).
Los que tienen seis o menos, o múltiplos de seis, ejes de simetría en su estructura corporal son denominados hexacorales (subclase Hexacorallia).Este grupo incluye a los principales corales constructores de arrecifes, los pertenecientes al orden Scleractinia (escleractinios). Los otros géneros de corales hermatípicos pertenecen a la subclase Octocorallia (como Heliopora y Tubipora), y a la clase Hydrozoa (como Millepora).
Los factores ecológicos necesarios para el crecimiento de los corales hermatípicos son, entre otros:
- Aguas relativamente someras (desde la superficie hasta varias decenas de metros).
- Temperaturas cálidas (entre 20 y 30 °C).
- Salinidades normales (entre 27 y 40 °C).
- Fuerte penetración de la luz.
- Intercambio con las aguas oceánicas abiertas en zonas agitadas de alta energía, lo que hace que se trate de aguas claras, sin sedimento en suspensión y con cierto contenido en nutrientes.
- Sustrato firme para su anclaje.
Solo en el Caribe, existen por lo menos 50 especies de corales pétreos, cada una con una estructura esquelética única.
Algunos tipos conocidos son:
- Corales cerebro, que pueden crecer hasta 1,8 metros de ancho.
- Acropora, que crecen rápidamente y pueden tener un tamaño grande; son importantes constructores de arrecifes. Especies como Acropora cervicornis tienen grandes ramas, algo semejantes a cuernos de ciervos, y habitan en zonas con un oleaje fuerte.
- Dendrogyra, que forman columnas que pueden llegar a tener una altura de 3 metros.
- Leptopsammia, que aparece en casi todas las partes del mar Caribe.
Corales ahermatípicos
Los corales ahermatípicos no construyen arrecifes, porque no generan un esqueleto. Tienen ocho tentáculos y se conocen también como octocorales, subclase Octocorallia. Incluyen los corales del orden Alcyonacea, así como algunas especies en el orden Antipatharia (coral negro, géneros Cirrhipathes y Antipathes). Corales ahermatípicos, como los gorgonáceos y Sessiliflorae, son también conocidos como corales blandos. A diferencia de los corales pétreos, son flexibles, ondulando en las corrientes de agua, y a menudo están perforados, con una apariencia de encaje.
Sus esqueletos son proteínicos en lugar de calcáreos. Los llamados corales blandos y corales cuero o piel, en su mayoría de aspecto carnoso, tienen en sus tejidos unos cristales microscópicos de calcita llamados espículas, cuya función es dar consistencia al tejido del animal, en ausencia de esqueleto propiamente dicho. La forma y distribución de las espículas son las principales características utilizadas en la identificación de géneros y especies de octocorales.Los corales blandos son un poco menos abundantes (en el Caribe ocurren 20 especies, frente a las 197 de corales duros) que los corales pétreos.
Corales porosos
Los corales pueden ser porosos o no porosos. Los primeros tienen esqueletos porosos que permiten que sus pólipos se conecten entre sí a través del esqueleto. Corales duros no porosos tienen esqueletos sólidos, macizos.
Características
El animal conocido como coral, el pólipo, mide desde apenas unos milímetros a algunos centímetros de diámetro. Los del orden Scleractinia tienen la capacidad de fijar sobre sus tejidos el carbonato de calcio disuelto en el mar y así formar las estructuras rígidas características.
La estructura calcárea del coral es blanca, los diferentes colores que presentan se deben, tanto a los diferentes pigmentos de sus tejidos, como en otros filos animales, como a unas microalgas que viven en simbiosis con la mayoría de los pólipos coralinos y que reciben el nombre de zooxantelas. Tan solo algunos géneros de corales como Tubastraea, Dendronephthya o parte de las gorgonias no son fotosintéticos. Las algas realizan la fotosíntesis produciendo oxígeno y azúcares, que son aprovechados por los pólipos, y se alimentan de los catabolitos del coral (especialmente fósforo y nitrógeno). Esto, en el caso de los corales fotosintéticos, les proporciona del 70 al 95% de sus necesidades alimenticias. El resto lo obtienen atrapando plancton. Por esta razón el coral necesita aguas transparentes para desarrollarse, para que las zooxantelas realicen así la fotosíntesis.
Los corales no fotosintéticos son animales omnívoros, y se alimentan tanto de zooplancton como de fitoplancton o materia orgánica disuelta en el agua.
Anatomía

En el siglo IX, el erudito musulmán Al-Biruni clasificó esponjas y corales como animales, argumentando que responden al tacto. Sin embargo, por lo general se creía que los corales eran plantas, hasta que, en el siglo XVIII, William Herschel utilizó un microscopio para determinar que los corales tenían las membranas celulares delgadas características de un animal.
Forma colonial
Los pólipos se interconectan mediante un tejido colonial común, denominado cenénquima, compuesto por mesoglea en la que se encuentran embebidos los pólipos, las escleritas, y los canales gastrodermales. Un sistema complejo y bien desarrollado de canales gastrovasculares, que se utilizan para repartir de manera significativa los nutrientes y simbiontes. En los corales blandos, estos canales varían en tamaño de 50-500 micrómetros (0,0050-0,050 cm) de diámetro, y permiten el transporte de metabolitos y componentes celulares.

Pólipo
Aunque una colonia de coral puede dar la impresión visual de un solo organismo, es en realidad un conjunto que se compone de muchos organismos multicelulares individuales, bien que genéticamente idénticos, conocidos como pólipos. Los pólipos tienen por lo general unos pocos milímetros de diámetro, y se componen de una capa externa de epitelio y tejido interno gelatinoso conocido como mesoglea. Son radialmente simétricos, con tentáculos que rodean una boca central, la única apertura hacia el estómago o celenteron, a través de la cual se ingieren los alimentos y se expulsan los residuos.
Exoesqueleto
El estómago se cierra en la base del pólipo, donde el epitelio produce un exoesqueleto llamado placa basal o calículo (L. pequeña taza). El calículo está formado por un anillo calcáreo engrosado (engrosamiento anular) con seis, o múltiplos de seis, crestas radiales de apoyo. Estas estructuras crecen verticalmente y se proyectan dentro de la base del pólipo. Cuando un pólipo está físicamente estresado, sus tentáculos se contraen en el cáliz de modo que prácticamente ninguna parte se expone por encima de la plataforma del esqueleto. Esto protege al organismo de los depredadores y los elementos.
El pólipo crece, por extensión de cálices verticales que de vez en cuando septan para formar una nueva placa basal superior. Tras muchas generaciones, estas extensiones constituyen las grandes estructuras calcáreas de los corales y finalmente los arrecifes coralinos.
La formación del exoesqueleto calcáreo involucra la deposición del mineral aragonito por los pólipos, sobre la base de calcio e iones de carbonato que adquieren a partir de agua de mar. La velocidad de deposición, que varia enormemente dependiendo de las especies y las condiciones ambientales, puede llegar a ser 10 g/m² de pólipo/día. Esto depende también de la luz, con la producción nocturna siendo 90% menor que la de mediodía. El exoesqueleto individual de cada pólipo se denomina coralito.

Tentáculos
Los nematocistos en las puntas de los tentáculos son células urticantes que llevan veneno, que se liberan rápidamente en respuesta al contacto con otro organismo. Los tentáculos también tienen una franja de contracción de epitelio llamado faringe. Las medusas y anémonas de mar también tienen nematocistos.

Alimentación
Los pólipos se alimentan de una variedad de pequeños organismos, desde plancton demersal microscópico hasta pequeños peces. Los tentáculos del pólipo inmovilizan o matan a sus presas con sus nematocistos (también conocido como "cnidocito"). Luego se contraen para dirigir la presa al estómago. Una vez digerida la presa, el estómago se vuelve a abrir, lo que permite la expulsión de desechos y el comienzo del siguiente ciclo de caza. Los pólipos también recogen moléculas orgánicas y moléculas orgánicas disueltas.
Zooxantelas simbiontes
Muchos corales, así como otros grupos de cnidarios tales como Aiptasia (un género de anémonas de mar), forman una relación simbiótica con una clase de algas, zooxantelas, del género Symbiodinium, un dinoflagelado. Aiptasia, una plaga conocida entre los aficionados de acuarios de coral debido a su sobrecrecimiento en la roca viva, sirven como un valioso organismo modelo en el estudio de la simbiosis cnidarios-algas. Típicamente, cada pólipo alberga una especie de alga. A través de la fotosíntesis, estos proporcionan energía al coral, y ayudan en la calcificación. Hasta un 30% del tejido de un pólipo puede ser material vegetal.
Reproducción

En cuanto a la reproducción, existen especies de reproducción sexual y reproducción asexual, y en muchas especies donde se dan ambas formas. Las células sexuales son expulsadas al mar todas a la vez, siguiendo señales como las fases lunares o las mareas. La fecundación suele ser externa, no obstante, algunas especies mantienen el óvulo en su interior (cavidad gastrovascular) y es allí donde son fecundados los huevos; y las puestas son tan numerosas que llegan a teñir las aguas.
Muchos huevos son devorados por los peces y otras especies marinas, pero son tantos que, aunque el porcentaje de supervivencia oscila entre el 18 y el 25%, según estudios de biología marina, los supervivientes garantizan la continuidad de las especies.
Los huevos una vez en el exterior, permanecen a la deriva arrastrados por las corrientes varios días, más tarde se forma una larva plánula que, tras deambular por la columna de agua marina, se adhiere al sustrato o rocas y comienza su metamorfósis hasta convertirse en pólipo y nuevo coral.
Reproducción asexual


Dentro de una colonia de coral, los pólipos genéticamente idénticos se reproducen asexualmente, ya sea a través de gemación (ciernes) o por división longitudinal o transversal; ambas se muestran en la foto de Orbicella annularis.
Gemación
"Gemación" consiste en separar un pólipo menor de un adulto. A medida que crece el nuevo pólipo, se forman las partes del cuerpo. La distancia entre el nuevo pólipo y el adulto crece, y con ella, el coenosarco (el cuerpo común de la colonia; véase anatomía de un pólipo). La gemación puede ser:
- Intratentacular: desde sus discos orales, produciendo pólipos del mismo tamaño dentro del anillo de tentáculos.
- Extratentacular: a partir de su base, produciendo un pólipo menor.
División
"División" forma dos pólipos, tan grandes como el original. "División longitudinal" comienza cuando un pólipo se ensancha y luego divide su celenteron, análogo a la división longitudinal de un tronco. La boca también se divide y forma nuevos tentáculos. Luego los dos "nuevos" pólipos generan las demás partes corporales y el exoesqueleto. "División transversal" se produce cuando los pólipos y el exoesqueleto se dividen transversalmente en dos partes. Esto significa que una parte tiene el disco basal (la parte inferior) y la otra tiene el disco oral (parte superior), semejante a cortar el extremo de un tronco. Los nuevos pólipos tienen que generar las piezas que faltan individualmente.
La reproducción asexual tiene varios beneficios para estos organismos coloniales sésiles:
- La clonación permite altas tasas de reproducción y una rápida explotación del hábitat.
- El crecimiento modular permite el aumento de la biomasa sin una disminución correspondiente en la relación superficie-volumen.
- El crecimiento modular retrasa la senescencia, al permitir que el clon puede sobrevivir la pérdida de uno o más módulos.
- Los nuevos módulos pueden sustituir los módulos muertos, reduciendo la mortalidad de los clones y preservando el territorio ocupado por la colonia.
- La difusión de clones a lugares distantes reduce la mortalidad entre clones causada por amenazas localizadas.
División de colonia
Colonias enteras pueden reproducirse asexualmente, formando dos colonias con el mismo genotipo.
- "Fisión" ocurre en algunos corales, en particular dentro de la familia Fungiidae, en la cual la colonia se divide en dos o más colonias durante las primeras etapas de desarrollo.
- "Abandono" se produce cuando un pólipo único abandona la colonia y se asienta sobre un sustrato diferente para crear una nueva colonia.
- "Fragmentación" involucra a pólipos individuales desglosados de la colonia durante tormentas u otros disturbios. Los pólipos separados pueden iniciar nuevas colonias.
Arrecifes de coral

Los pólipos de coral mueren con el tiempo, pero las estructuras calcáreas se mantienen y pueden ser colonizadas por otros pólipos de coral, que seguirán creando estructuras cálcicas generación tras generación. A lo largo de miles o de millones de años se forman grandes estructuras calcáreas conocidas como arrecifes de coral.
En ocasiones los arrecifes son tan grandes que pueden llegar a emerger de la superficie. Así, cuando el coral crece alrededor de una isla volcánica que posteriormente se hunde, se crea una estructura coralina en forma de anillo con una laguna central que recibe el nombre de atolón.
El arrecife de mayor longitud es la Gran Barrera de Arrecifes, en la costa de Queensland en Australia: tiene más de 2000 km², y es una de las construcciones naturales más grandes del mundo. La región del mundo con más especies de corales y más biodiversidad en sus arrecifes coralinos es el Triángulo de coral, en el sureste asiático, que incluye más de 500 especies de corales (el 76% de las especies coralinas conocidas) y al menos 2228 especies de peces.
Los arrecifes coralinos forman el hogar de muchos organismos marinos que allí encuentran alimento y protección contra los depredadores.
El segundo arrecife coralino más grande del mundo, el Arrecife Mesoamericano (a lo largo de la costa de México, Belice, Guatemala y Honduras), se encuentra en el mar Caribe, y se extiende por más de 700 km desde la península de Yucatán hasta las Islas de la Bahía en la costa norte de Honduras. Aun cuando mide un tercio de lo que mide la Gran Barrera Arrecifal de Australia, el Arrecife del Caribe Mesoamericano alberga una gran diversidad de organismos, incluidos 60 tipos de corales y más de 500 especies de peces.
El ecosistema también es el sitio de dos grandes iniciativas internacionales de conservación, una ya bien establecida y otra apenas en sus inicios.
En 1998, el Fondo Mundial para la Naturaleza (WWF, por sus siglas en inglés) identificó al arrecife del Caribe mesoamericano como un ecosistema prioritario y una ecorregión de importancia global, por lo que comenzó un esfuerzo de conservación del arrecife a largo plazo.
Historia evolutiva

Aunque los corales aparecieron por primera vez durante el Cámbrico, hace 542 millones de años, los fósiles correspondientes son extremadamente raros hasta el período Ordovícico, 100 millones de años más tarde, cuando se generalizó la distribución de los corales rugosos y tabulados.
El registro fósil de los corales tabulados se halla en piedra caliza y lutita calcáreas (en inglés: shale) del Ordovícico y Silúrico y con frecuencia forman cojines bajos o ramificaciones junto a los corales rugosos. Su número comenzó a declinar a mediados del Silúrico, y se extinguieron a finales del Pérmico, hace 250 millones de años. Los esqueletos de los corales tabulados se componen de una forma de carbonato de calcio conocido como calcita.
Los corales rugosos llegaron a ser dominantes a mediados del Silúrico, y se extinguieron a principios del Triásico. Los corales rugosos vivieron solitarios y en colonias, y sus exoesqueletos se componían también de calcita.
Los corales escleractinios llenaron el vació dejado por las especies de corales rugosos y tabulados extintas. Sus fósiles pueden encontrarse en pequeñas cantidades en las rocas del período Triásico, y llegaron a ser comunes desde el Jurásico. Los esqueletos de los escleractinios se componen de un tipo de carbonato de calcio conocido como aragonita. A pesar de que son geológicamente más jóvenes que los corales tabulados y rugosos, su registro fósil es menos completo debido a su esqueleto de aragonita, cuya conservación es más difícil.
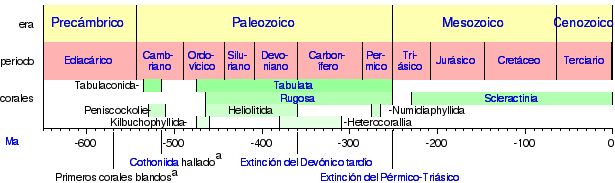 |
|
|
Cronograma del registro fósil coralino y de los principales acontecimientos desde hace 650 Ma hasta el presente. |
|
Los corales eran muy abundantes en ciertas épocas del pasado geológico. Al igual que los corales modernos, estos antepasados coralinos construyeron arrecifes, algunos de los cuales terminaron como grandes estructuras en rocas sedimentarias.
Los fósiles de otros habitantes arrecifales, como algas, esponjas, y los restos de muchos equinodermos, braquiópodos, bivalvos, gasterópodos, y trilobites, aparecen junto con los fósiles de coral. La distribución de los fósiles de coral no está limitada a los restos de arrecifes, ya que se pueden encontrar muchos fósiles solitarios en otros lugares, tales como Cyclocyathus que ocurre en la formación de arcilla de Gault en Inglaterra.
Estado de conservación
Amenazas


Según algunos estudios, los arrecifes de coral están en descenso en el mundo entero. Las principales amenazas localizadas para los ecosistemas coralinos son la extracción de coral, la escorrentía agrícola y urbana, la contaminación de orgánicos e inorgánicos, la sobrepesca, la pesca con explosivos, las enfermedades coralinas y la excavación de canales de acceso a islas y bahías. Las amenazas más amplias incluyen el aumento de la temperatura del mar, la subida del nivel del mar, y el cambio del pH debido a la acidificación de los océanos, todos asociados con las emisiones de gases de efecto invernadero. En 1998, el 16% del total de arrecifes de coral murieron como consecuencia del aumento de la temperatura del mar.
Cambios de temperatura del agua de más de 1-2 grados Celsius o cambios de salinidad, pueden diezmar los corales. Bajo tales presiones ambientales, los corales expulsan sus zooxantelas; sin ellos los tejidos del coral revelan el blanco de sus esqueletos, un evento conocido como blanqueo de coral.
Estimaciones globales indican que aproximadamente el 10% del total de los arrecifes de coral está muerto. Alrededor del 60% de los arrecifes coralinos está en riesgo como resultado de actividades humanas. Se estima que la destrucción de los arrecifes coralinos puede llegar al 50% en el año 2030. En respuesta, la mayoría de las naciones establecieron leyes ambientales en un intento de proteger este importante ecosistema marino.
Entre el 40% y el 70% de las algas comunes, transfieren metabolitos lípidosolubles y causan decoloración y muerte entre los corales, particularmente cuando se da una sobrepoblación de algas. Las algas proliferan cuando tienen suficientes nutrientes como resultado de contaminación orgánica, y si la sobrepesca dramáticamente reduce el pastoreo por herbívoros, como el pez loro.
Protección

Los arrecifes de coral pueden ser protegidos de daños antropogénicos si son declarados zonas protegidas, por ejemplo área marina protegida, reserva de la biosfera, parque marino, monumento nacional, patrimonio de la humanidad, gestión de la pesca y protección de hábitat.
Muchos gobiernos ahora prohíben la extracción de coral de los arrecifes e informan a los residentes de la costa sobre su protección y su ecología. Aunque medidas locales, como la protección y restauración del hábitat de herbívoros marinos, puede reducir los daños locales, las amenazas globales y más a largo plazo, como la acidificación, el cambio de temperatura y el aumento del nivel del mar, siguen siendo un desafío. De hecho, recientes estudios han constatado que en 2016 ha muerto aproximadamente el 35% de los corales en 84 áreas de las secciones norte y centro de la Gran Barrera de Coral, debido al blanqueo de coral producido por el aumento de la temperatura del mar.
Algunos géneros y especies de coral
-

Especies de Acropora.
-

Antipathes dichotoma o coral negro.
-

Corallium rubrum o coral rojo.
-

Dendronephthya, coral blando.
-

Coral cerebro de la familia Faviidae.
-

Fungia scutaria o coral hongo.
-

Gorgonia flabellum o gorgonia de Venus.
-

Heliopora coerulea o coral azul.
-

Millepora alcicornis o coral de fuego.
-

Sarcophyton o coral cuero con pólipos retraidos.



.jpg)




.jpg)

Véase también
 En inglés: Coral Facts for Kids
En inglés: Coral Facts for Kids
