
Biota del periodo Ediacárico para niños
Los organismos ediacáricos son formas de vida muy antiguas que vivieron en la Tierra hace entre 635 y 542 millones de años, durante un periodo llamado Ediacárico. Estos fueron los primeros organismos multicelulares complejos que conocemos, es decir, estaban formados por muchas células y eran lo suficientemente grandes como para verlos sin microscopio.
Aparecieron poco después de una gran glaciación que cubrió gran parte del planeta. La mayoría de ellos desaparecieron justo antes de un evento llamado la explosión cámbrica, que fue un momento en el que la vida en la Tierra se diversificó muy rápidamente y aparecieron muchos de los grupos de animales que conocemos hoy. Pocos organismos ediacáricos se parecen a los animales modernos, y las nuevas formas de vida del Cámbrico los reemplazaron por completo.
Los organismos ediacáricos surgieron hace unos 580 millones de años y prosperaron hasta hace 542 millones de años. Aunque se han encontrado algunos fósiles que podrían ser supervivientes en épocas posteriores, la mayoría de estas comunidades desaparecieron al final del periodo Ediacárico. Hay varias ideas sobre por qué desaparecieron, como cambios en el ambiente, la aparición de nuevos depredadores o la competencia con otras formas de vida.
La forma en que estos organismos se relacionan con la vida actual es un gran misterio para los científicos. Algunos parecen estar relacionados con grupos que hoy son comunes, como las esponjas o las medusas, o incluso los moluscos. Sin embargo, la mayoría de los fósiles de gran tamaño son muy diferentes a las formas de vida posteriores: parecen discos, bolsas o colchones blandos. Por eso, algunos paleontólogos creen que los organismos ediacáricos podrían haber sido una rama de la vida que se extinguió por completo, un "experimento fallido" de la vida multicelular.
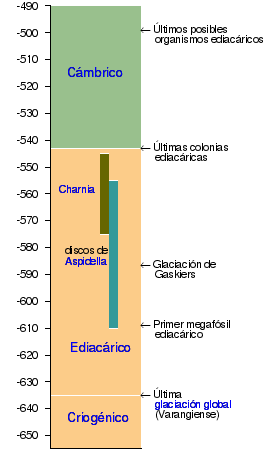
| Neoproterozoico (última era del Precámbrico) |
| Paleozoico (primera era del Fanerozoico) |
Escala del eje: hace millones de años.
Referencias: Waggoner 1998, Hofmann 1990
Contenido
¿Cómo se descubrieron los organismos ediacáricos?
Los primeros fósiles ediacáricos se encontraron en 1868. Eran unos discos llamados Aspidella terranovica. Su descubridor, Alexander Murray, pensó que eran útiles para saber la edad de las rocas en Terranova. Sin embargo, como estaban en rocas más antiguas que las del Cámbrico (que se creía que tenían la primera vida), pasaron años hasta que alguien sugirió que podían ser fósiles. Al principio, la idea fue rechazada y se pensó que eran formaciones de gases o rocas sin vida.
En 1933, se encontraron fósiles parecidos en Namibia, pero también se creyó que eran del Cámbrico. En 1946, Reg Sprigg descubrió fósiles con forma de medusa en los montes Ediacara en Australia. Aunque interesantes, no llamaron mucho la atención porque se pensaba que esas rocas también eran del Cámbrico.

No fue hasta 1957, con el descubrimiento de Charnia en Inglaterra, que los científicos empezaron a tomar en serio la idea de que había vida multicelular antes del Cámbrico. Este fósil, con forma de hoja, se encontró en rocas que sin duda eran del Precámbrico. El paleontólogo Martin Glaessner conectó este hallazgo con los anteriores, y con mejores métodos de datación, se descubrieron muchos más organismos ediacáricos.
Al principio, los fósiles se encontraban en rocas muy rugosas que no conservaban bien los detalles. Pero en 1967, S. B. Misra encontró fósiles en capas de ceniza volcánica en Terranova y Labrador. Esta ceniza fina permitió ver detalles que antes no se podían, lo que ayudó mucho a entender cómo eran estos organismos.
Debido a la dificultad para comparar rocas de diferentes partes del mundo, se usaron muchos nombres para estos organismos. Finalmente, en 2004, la Unión Internacional de Ciencias Geológicas decidió llamar a este periodo el "Ediacárico" en honor a los montes Ediacara en Australia.
¿Cómo se conservaron estos organismos?
La mayoría de los fósiles que encontramos son de partes duras, como huesos o conchas. Por eso, es sorprendente que los organismos ediacáricos, que tenían cuerpos blandos y no tenían esqueleto, se hayan conservado tan bien. Una razón podría ser que no había muchas criaturas que excavaran en el fondo marino en ese tiempo. Más tarde, en el Cámbrico, cuando aparecieron animales excavadores, las huellas de cuerpos blandos se destruían antes de poder fosilizarse.
Estromatolitos y su papel en la conservación


Los estromatolitos son estructuras formadas por colonias de microorganismos que crean "tapetes" pegajosos. Estos tapetes atrapan partículas de sedimento y crecen hacia arriba. Si una capa de sedimento los cubre demasiado rápido, algunas partes de la colonia mueren, dejando fósiles con una textura arrugada que parece "piel de elefante".
Muchos fósiles ediacáricos se encuentran en superficies con esta textura de estromatolitos. Aunque los estromatolitos eran muy comunes en el pasado, su número disminuyó mucho en el Cámbrico debido a la aparición de animales que se alimentaban de ellos. Hoy en día, solo se encuentran en lugares donde estos animales no pueden sobrevivir.
El proceso de fosilización
La conservación de estos fósiles es un misterio fascinante, ya que los organismos de cuerpo blando no suelen fosilizarse. A diferencia de otros fósiles de cuerpo blando que se encuentran en lugares específicos con condiciones raras, los organismos ediacáricos se han encontrado en todo el mundo. Esto sugiere que algo muy diferente ocurría en el periodo Ediacárico que permitía su conservación.
La teoría más aceptada es que estos fósiles se conservaron porque fueron cubiertos rápidamente por ceniza o arena, lo que los atrapó junto al barro o los tapetes de microbios donde vivían. Las capas de ceniza son especialmente útiles porque conservan muchos detalles y se pueden fechar con mucha precisión.
También es común encontrar fósiles ediacáricos en capas de arena depositadas por tormentas o corrientes oceánicas fuertes. Hoy en día, los organismos de cuerpo blando casi nunca fosilizan así. Sin embargo, la presencia de grandes capas de microbios probablemente ayudó a preservar las huellas de los organismos ediacáricos al estabilizar el sedimento.
¿Qué partes se conservaron?
La forma en que se conservó un organismo dependía de la rapidez con la que el sedimento lo cubría y de la rapidez con la que se descomponía. La mayoría de los fósiles con forma de disco se descomponían antes de ser cubiertos por completo, dejando un molde de su parte inferior. En cambio, los organismos esponjosos solían descomponerse después de que el sedimento los cubriera, por lo que se conservaba su superficie superior. Estos organismos esponjosos eran más resistentes y a veces se encuentran incluso dentro de capas de sedimento formadas por tormentas. En algunos casos, las bacterias ayudaron a formar un "molde" mineral del organismo.
¿Cómo eran los organismos ediacáricos?
Datos para niños
|
|
|---|---|
| Tateana inflata (Cyclomedusa radiata), un fósil en forma de disco que formó parte de algún animal desconocido. Escala métrica. |  |
| Un molde del esponjoso Charnia, el primer organismo precámbrico complejo conocido. Antiguamente se lo consideraba pariente de los Pennatulacea. |  |
| Spriggina, un posible precursor de los trilobites, podría haber sido uno de los depredadores que causó la extinción de los organismos ediacáricos y la posterior diversificación de los animales. |  |
| Un icnofósil de finales del Ediacárico, preservado en la base de un estrato. |  |
| Cadena de icnofósiles creados por la especie Yorgia, un organismo que se alimentaba del fondo, junto al fósil del propio cuerpo del organismo (derecha). |  |
Los organismos ediacáricos tenían una gran variedad de formas. Su tamaño iba desde unos pocos milímetros hasta varios metros. Su complejidad variaba desde simples "manchas" hasta estructuras muy elaboradas. Podían ser rígidos o blandos como las medusas. Tenían casi todos los tipos de simetría. Se diferenciaban de los fósiles anteriores por su estructura multicelular organizada y su tamaño considerable.
Podemos agrupar la variedad de formas de los organismos ediacáricos en varios tipos: embriones, discos, bolsas, organismos esponjosos, y huellas fósiles (icnofósiles).
Fósiles con forma de embriones
Se han encontrado fósiles con forma de embriónes en rocas precámbricas, especialmente en China. Algunos de estos hallazgos generaron mucho interés. Sin embargo, algunos científicos creen que en realidad son estructuras no orgánicas formadas por minerales. Otros "embriones" se han interpretado como restos de bacterias gigantes.
Microfósiles de hace 632.5 millones de años podrían ser "etapas de reposo" de los animales más antiguos conocidos. Otra idea es que estas estructuras son las formas adultas de los animales de ese periodo. Estudios recientes sugieren que no son embriones de animales, sino acumulaciones de células similares a quistes de protozoos, que usaban estas estructuras para reproducirse.
Fósiles con forma de discos
Fósiles circulares como Ediacaria, Cyclomedusa y Rugoconites hicieron que al principio se pensara que los organismos ediacáricos eran cnidarios, el grupo que incluye a las medusas y los corales.
Sin embargo, estudios posteriores han llevado a otras ideas. Ahora, ninguno de estos fósiles se considera con certeza una medusa. Otras interpretaciones sugieren que podrían ser protistas o anémonas marinas. Algunos patrones sugieren que eran colonias de microbios. Como solo se ha fosilizado la parte inferior de estos organismos, no se conocen suficientes detalles para entender bien su estructura.
Fósiles con forma de bolsas
Fósiles como Pteridinium, que se conservan dentro de capas de sedimento, parecen "bolsas llenas de barro". Los científicos aún no se ponen de acuerdo sobre cómo interpretarlos.
Organismos esponjosos
Los organismos que se parecen a un colchón inflable son un grupo importante. A veces, estas "esponjas" se rompían antes de fosilizarse, y estos ejemplares dañados dan pistas valiosas sobre cómo eran. Por ejemplo, las hojas de Swartpuntia germsi solo se ven en ejemplares rotos, porque normalmente quedaban ocultas al ser aplastados.
Estos organismos parecen dividirse en dos grupos: los "rangeomorfos" con formas repetitivas (fractales) y los "erniettomorfos", más simples. Incluyen a los famosos Charnia y Swartpuntia. Son los más característicos de los organismos ediacáricos, pero también los más difíciles de ubicar en el árbol filogenético. Como no tenían boca, intestinos, ni órganos reproductivos visibles, su forma de vida es muy extraña para nosotros. La idea más aceptada es que absorbían los nutrientes del agua a su alrededor por ósmosis.
Organismos ediacáricos "no ediacáricos"
Algunos organismos ediacáricos han conservado detalles más complejos, lo que ha permitido interpretarlos como posibles formas primitivas de grupos de animales que existen hoy.
El fósil más antiguo de este tipo es Vernanimalcula, que tiene simetría bilateral (dos lados iguales). Ejemplos más recientes, aceptados como bilaterales, incluyen Kimberella, parecido a un molusco, Spriggina, y Parvancorina, con forma de escudo. Sus relaciones con los animales modernos aún se debaten.
También se han encontrado pequeños fósiles con caparazón en el periodo Ediacárico, como Cloudina, un pequeño tubo que muestra marcas de depredación. Esto sugiere que, aunque los depredadores no eran muy comunes, ya existían.
En el periodo Ediacárico ya había representantes de grupos modernos, como esponjas, algas rojas y verdes, protistas y bacterias. Algunos de ellos ya existían mucho antes. También se han descrito posibles artrópodos.
Icnofósiles (huellas y rastros)
Con algunas excepciones, las únicas cuevas o túneles conocidos del periodo Ediacárico son horizontales, en la superficie o justo debajo. Estas huellas sugieren que existían organismos móviles con cabeza, que probablemente tenían simetría bilateral. Sin embargo, también podrían ser el resultado del movimiento de organismos más simples que rodaban mientras se alimentaban en el fondo marino.
Algunos fósiles ediacáricos, especialmente los discos, se interpretaron al principio como huellas, pero esta idea no se confirmó. Además de los túneles, se han encontrado huellas directamente asociadas con un fósil ediacárico. Yorgia y Dickinsonia a menudo aparecen al final de largos rastros que coinciden con su forma o con su forma de alimentarse. El posible molusco Kimberella está asociado con marcas de raspado, quizás hechas por una rádula (una especie de lengua con dientes).
¿De dónde vinieron?
Cuando aparecieron los organismos ediacáricos, hace 655 millones de años, la Tierra ya tenía casi 4000 millones de años. Aunque hay fósiles de hace 3460 millones de años, la primera prueba clara de vida es de hace 2700 millones de años, y hace 1200 millones de años ya existían células con núcleo. Entonces, ¿por qué tardó tanto en aparecer vida compleja como la ediacárica?
Una explicación es que la evolución es un proceso lento que simplemente necesitó 4000 millones de años para que se acumularan las adaptaciones necesarias. De hecho, parece que hubo un aumento gradual en la complejidad de la vida a lo largo del tiempo, con formas cada vez más complejas.

Otra idea es que no era ventajoso ser grande hasta que aparecieron los organismos ediacáricos. El ambiente favorecía a las formas pequeñas, como el plancton actual, que se reproduce rápido para aprovechar los nutrientes. Sin embargo, el ambiente tendría que haber sido muy diferente para que un tamaño mayor nunca fuera una ventaja.
Uno de los factores principales que limitan el tamaño de los organismos es la cantidad de oxígeno en la atmósfera. Sin un sistema circulatorio complejo, las bajas concentraciones de oxígeno hacen muy difícil que un organismo grande lo absorba lo suficientemente rápido para sus necesidades.
En la Tierra primitiva, el oxígeno reaccionaba con elementos como el hierro. El oxígeno no pudo acumularse en la atmósfera hasta que todo el hierro se oxidó. Se han encontrado pruebas de cantidades significativas de oxígeno en la atmósfera justo antes de la aparición de los primeros fósiles ediacáricos. Esto sugiere que el oxígeno pudo haber sido clave para la diversificación de la vida en el Ediacárico. Parece que el oxígeno se acumuló en dos etapas: la aparición de pequeños organismos fijos coincidió con un primer aumento de oxígeno, y la aparición de organismos más grandes y móviles con un segundo.
Los periodos de frío intenso también se han sugerido como un obstáculo para la evolución de la vida multicelular. Los embriones más antiguos conocidos aparecen solo un millón de años después de una glaciación global, lo que sugiere que el hielo y los océanos fríos pudieron haber impedido la aparición de vida compleja. Es posible que la vida multicelular hubiera evolucionado antes de estas glaciaciones y que estas la extinguieran. Sin embargo, la diversidad de vida en la Antártida actual hace dudar si las temperaturas frías realmente frenan la evolución.
A principios de 2008, un equipo de científicos analizó la diversidad de organismos ediacáricos de tres lugares diferentes: Avalon (Canadá, hace 575-565 millones de años), mar Blanco (Rusia, 560-550 millones de años) y Nama (Namibia, 550-542 millones de años). Concluyeron que, aunque las muestras del mar Blanco tenían más especies, no había grandes diferencias entre los tres grupos. Esto sugiere que antes del periodo de Avalon, estos organismos debieron tener su propia "explosión" de diversidad, similar a la explosión del Cámbrico.
¿Por qué desaparecieron?
La falta de muchos fósiles hace que la desaparición de los organismos ediacáricos sea un misterio. Parece que hubo una desaparición bastante repentina al final del periodo Ediacárico. La presencia de organismos "ediacáricos" en el Cámbrico no es aceptada por todos los científicos. La causa de esta desaparición aún se debate.
Sesgo de conservación
La desaparición repentina de fósiles ediacáricos cerca del límite del Cámbrico podría deberse simplemente a que las condiciones ya no eran favorables para su fosilización. Podrían haber seguido viviendo sin dejar fósiles. Sin embargo, si hubieran sido comunes, se deberían haber encontrado más ejemplares en yacimientos fósiles excepcionalmente bien conservados, a menos que estos lugares no fueran adecuados para ellos o para su conservación.

Depredación y organismos que se alimentaban del fondo
Se ha sugerido que, en el Cámbrico, los organismos que estaban más arriba en la cadena trófica hicieron que las capas de microbios casi desaparecieran. Si estos organismos que se alimentaban del fondo aparecieron al mismo tiempo que los organismos ediacáricos empezaron a disminuir, podría ser que desestabilizaran el fondo marino, causando al menos un cambio en las comunidades ediacáricas o incluso su extinción.
Otra posibilidad es que los nuevos animales con esqueleto se alimentaran directamente de los organismos ediacáricos, que eran relativamente indefensos. Sin embargo, si la interpretación de Kimberella (un organismo ediacárico) como un "organismo que se alimentaba del fondo" es correcta, esto sugiere que los ediacáricos ya enfrentaban una amenaza limitada de depredación. Además, hay pocas huellas fósiles de depredación en el periodo Ediacárico, lo que va en contra de la teoría de los depredadores activos.

Competencia
Es posible que un aumento de la competencia, debido a la evolución de nuevas características en otros grupos, expulsara a los organismos ediacáricos de sus hábitats. Sin embargo, este argumento no ha explicado bien fenómenos similares en otros momentos de la historia de la vida.
Cambio en las condiciones ambientales
Aunque es difícil saber el efecto exacto de los cambios planetarios en los organismos, hubo grandes cambios al final del Precámbrico y principios del Cámbrico. La separación de los supercontinentes, el aumento del nivel del mar (creando mares poco profundos y "aptos para la vida"), la escasez de nutrientes, los cambios en la atmósfera (incluyendo los niveles de oxígeno y dióxido de carbono), y los cambios en la química de los océanos que favorecieron la formación de minerales, son factores que pudieron haber contribuido a la extinción de la vida ediacárica.
Tipos de comunidades ediacáricas
Se han encontrado fósiles de tipo Ediacara en veinticinco lugares de todo el mundo y en diferentes condiciones. Por eso, se agrupan en tres tipos principales de comunidades, que llevan el nombre de los lugares donde se encontraron por primera vez. Cada tipo de comunidad tiende a ocupar su propia región y, después de una primera diversificación, no cambia mucho durante el resto de su existencia.
Comunidad tipo Avalon
La comunidad tipo Avalon se define por el yacimiento de Mistaken Point, en Canadá. Es fácil fecharla porque contiene muchas capas finas de ceniza que son buenas para la datación radiométrica. Estas capas finas también conservan detalles muy delicados. Los organismos de esta comunidad parecen haber sobrevivido hasta la extinción de todos los organismos ediacáricos al comienzo del Cámbrico.
|
|
|---|
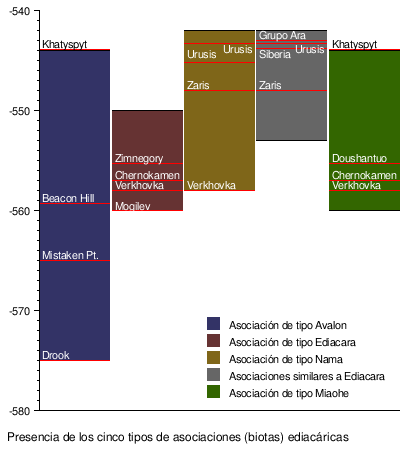 |
| Escala del eje: hace millones de años, datación por U/Pb de zircones. |
Estos organismos incluyen a los rangeomorfos de las profundidades marinas, como Charnia, que tienen un patrón de crecimiento repetitivo. Es probable que se conservaran en el lugar donde murieron. Aunque esta comunidad es menos diversa que las de tipo Ediacara o Nama, se parece a las comunidades de alimentación por filtración del periodo Carbonífero. Esto sugiere que los organismos se alimentaban filtrando el agua, ya que vivían en aguas demasiado profundas para la fotosíntesis. La baja diversidad podría deberse a la profundidad del agua o a que la comunidad era muy joven.
Comunidad tipo Ediacara
La comunidad tipo Ediacara lleva el nombre de las colinas Ediacara en Australia. Consiste en fósiles conservados cerca de las desembocaduras de ríos. Se encuentran en capas de arena y limo formadas bajo el agua, pero lo suficientemente poco profundas como para ser afectadas por las olas durante las tormentas. La mayoría de los fósiles se conservan como huellas en capas microbianas, pero algunos están dentro de capas de arena.
Comunidad tipo Nama
La comunidad tipo Nama se define por el yacimiento homónimo en Namibia. El tipo de fósiles más común tiene una conservación tridimensional, con organismos conservados en capas de arena con estructuras internas. Algunos científicos creen que estas formas eran organismos que excavaban, mientras que otros piensan que vivían en la superficie. Estas capas se encuentran entre areniscas, limos y esquistos, y a menudo contienen fósiles. Se cree que el ambiente eran barreras de arena formadas en la boca de los ríos.
¿Qué nos dicen estas comunidades?
En la región del mar Blanco de Rusia, se han encontrado los tres tipos de comunidades muy cerca unos de otros. Esto, junto con el hecho de que los organismos se solapan en el tiempo, hace que sea poco probable que representen etapas evolutivas o comunidades diferentes en el tiempo. Como se encuentran en todo el mundo (en todos los continentes excepto la Antártida), los límites geográficos no parecen influir. Se encuentran los mismos fósiles en todas las latitudes antiguas y en diferentes cuencas de sedimentos.
Lo más probable es que las tres comunidades representen organismos adaptados a vivir en diferentes ambientes. Cualquier patrón aparente en la diversidad o la edad podría ser solo una impresión debido a la escasez de muestras descubiertas, ya que los yacimientos fósiles ediacáricos son muy raros. Como los organismos ediacáricos representan una etapa temprana en la historia de la vida multicelular, no es sorprendente que no ocuparan todos los modos de vida posibles. Se ha calculado que, de noventa y dos modos de vida posibles (combinaciones de cómo se alimentaban, su posición en la cadena alimentaria, su movilidad, etc.), solo una docena estaban ocupados al final del Ediacárico. En la comunidad tipo Avalon, solo se encuentran cuatro. La falta de depredación a gran escala y la excavación vertical fueron probablemente los factores más importantes que limitaron la biodiversidad ecológica. La aparición de estas actividades durante el Cámbrico permitió que el número de modos de vida ocupados aumentara a treinta.
Galería de imágenes
-

Dickinsonia costata, un organismo ediacárico icónico, que muestra la apariencia «acolchada» típica de muchos organismos de esta biota.

Véase también
 En inglés: Ediacaran biota Facts for Kids
En inglés: Ediacaran biota Facts for Kids
- Anexo:Géneros de la biota de Ediacara.
- Origen de la vida.
- Explosión de Avalon
- Explosión cámbrica
