
Sistema inmunitario para niños
El sistema inmunitario es como el equipo de seguridad de tu cuerpo. Está formado por muchas partes, como células especiales y sustancias químicas, que trabajan juntas para protegerte de cosas que pueden hacerte daño. Esto incluye desde pequeños gérmenes como bacterias y virus, hasta problemas internos como células que no funcionan bien. Su trabajo principal es reconocer lo que es peligroso y atacarlo para mantener tu cuerpo sano y en equilibrio.
Este sistema de defensa está en todo tu cuerpo. Algunas de sus partes son moléculas que flotan en la sangre y la linfa (un líquido parecido a la sangre), como los anticuerpos. Otras partes son células que se encuentran en órganos como la médula ósea, el timo, el bazo y los ganglios linfáticos. En la médula ósea se producen los leucocitos (también conocidos como glóbulos blancos), que son células especializadas en la defensa. Hay muchos tipos de leucocitos, como los neutrófilos, linfocitos, macrófagos y células dendríticas, que viajan por la sangre y el sistema linfático para llegar a donde se les necesita.
Hay dos formas principales en que el sistema inmunitario te protege:
- La inmunidad innata (o natural): Es la primera línea de defensa. Actúa de forma rápida y general contra cualquier invasor. Es como una alarma que suena de inmediato.
- La inmunidad adquirida (o adaptativa): Es más específica y aprende a reconocer a los invasores. Es como un equipo de detectives que recuerda a los criminales para atraparlos más rápido la próxima vez. Esta es la base de las vacunas.
Cuando el sistema inmunitario no funciona bien, pueden aparecer enfermedades. Si es menos activo de lo normal, se llama inmunodeficiencia, y el cuerpo es más propenso a infecciones. Si es demasiado activo y ataca al propio cuerpo, se llama autoinmunidad. La inmunología es la ciencia que estudia todo esto.
Datos para niños Sistema inmunitario |
||
|---|---|---|
 En la imagen, neutrófilos (en color amarillo) fagocitando bacterias del carbunco (en color naranja). Imagen obtenida mediante un microscopio electrónico de barrido. La línea blanca en la parte inferior izquierda equivale a 5 micrómetros.
|
||
| Estudiado (a) por | inmunología | |
| Información fisiológica | ||
| Función | Protección de un organismo ante agentes externos. | |
| Estructuras principales | ||
| Leucocitos o glóbulos blancos | ||
|
|
||
Contenido
- ¿Qué es el Sistema Inmunitario?
- Órganos del Sistema Inmunitario
- Barreras de Defensa del Cuerpo
- Inmunidad Adaptativa o Adquirida
- Problemas del Sistema Inmunitario
- Manipulación del Sistema Inmunitario en Medicina
- Cómo los Gérmenes Engañan al Sistema Inmunitario
- Historia de la Inmunología
- Galería de imágenes
- Véase también
¿Qué es el Sistema Inmunitario?
El sistema inmunitario es el conjunto de partes y procesos biológicos dentro de un ser vivo que le permiten mantener su equilibrio interno. Esto lo logra defendiéndose de ataques externos, como gérmenes (bacterias, virus), o de problemas internos, como células que crecen de forma descontrolada. Su función principal es reconocer lo que es dañino y reaccionar para proteger el cuerpo.
Componentes del Sistema Inmunitario
El sistema inmunitario está compuesto por:
- Moléculas solubles: Son proteínas y otras sustancias que se encuentran en líquidos como la sangre y la linfa. Ejemplos son los anticuerpos y las proteínas del sistema del complemento.
- Células: Son los glóbulos blancos especializados en la defensa. Se producen en la médula ósea y se mueven por la sangre y el sistema linfático. Algunos tipos importantes son:
* Neutrófilos * Linfocitos * Eosinófilos * Basófilos * Mastocitos * Monocitos * Células dendríticas * Macrófagos
Niveles de Defensa Inmunitaria
El cuerpo tiene diferentes niveles de defensa:
- Inmunidad innata: Es la defensa natural y rápida. Está presente en casi todos los seres vivos, desde bacterias hasta mamíferos. Reconoce señales de "peligro" o de la presencia de gérmenes.
- Inmunidad adquirida: Es una defensa más avanzada que tienen los vertebrados, como los humanos. Se adapta con el tiempo para reconocer gérmenes específicos de forma más eficiente. Crea una "memoria" para que, si el mismo germen vuelve, la respuesta sea más rápida y fuerte. Este proceso es la base de las vacunas.

Órganos del Sistema Inmunitario
El sistema inmunitario tiene órganos que se clasifican en primarios y secundarios:
- Órganos primarios: Son la médula ósea y el timo. Aquí es donde las células de defensa, especialmente los linfocitos, maduran y se preparan para su trabajo.
- Órganos secundarios: Incluyen los ganglios linfáticos, el bazo y el tejido linfoide asociado a las mucosas. En estos lugares, las células inmunitarias se encuentran con los gérmenes o sus partes (antígenos) y aprenden a atacarlos.
Barreras de Defensa del Cuerpo
El sistema inmunitario protege el cuerpo con varias líneas de defensa:
Primera Línea: Barreras Físicas y Químicas
La primera defensa son las barreras que impiden que los gérmenes entren al cuerpo:
- Barreras físicas: La piel es una barrera protectora. Las superficies internas, como las del tracto gastrointestinal y el respiratorio, también tienen mucosas que atrapan a los gérmenes. La tos, los estornudos, las lágrimas y la orina ayudan a expulsar los gérmenes mecánicamente.
- Barreras químicas: La piel y las vías respiratorias producen sustancias que matan microbios. La saliva, las lágrimas y la leche materna contienen enzimas que atacan bacterias. El ácido gástrico en el estómago destruye muchos gérmenes que ingerimos.
Además, en el intestino y otras partes del cuerpo, hay bacterias "buenas" (microbiota) que compiten con las bacterias dañinas por alimento y espacio, ayudando a mantener el equilibrio.
Segunda Línea: Inmunidad Innata
Si un germen logra pasar las barreras, la inmunidad innata actúa de inmediato. Esta respuesta no es específica, lo que significa que ataca a los gérmenes de forma general.
Fiebre
La fiebre es un aumento de la temperatura corporal que ayuda al sistema inmunitario a combatir infecciones. Una temperatura más alta puede hacer que los glóbulos blancos se muevan y ataquen mejor a los invasores.
Inflamación
La inflamación es una de las primeras respuestas a una infección. Los síntomas son enrojecimiento, hinchazón y calor. Esto ocurre porque aumenta el flujo de sangre a la zona afectada y las células inmunitarias se acumulan allí para liberar sustancias que atacan al germen y ayudan a curar el tejido dañado.
Sistema del Complemento
El sistema del complemento es un grupo de proteínas que trabajan juntas para atacar las superficies de las células extrañas. Pueden marcar a los gérmenes para que otras células los destruyan, o incluso matarlos directamente.
Tercera Línea: Células de la Inmunidad Innata
Los leucocitos (glóbulos blancos) son el segundo brazo de la inmunidad innata. Identifican y eliminan gérmenes:
- Fagocitos: Como los macrófagos y neutrófilos, "comen" a los gérmenes rodeándolos y digiriéndolos.
- Células dendríticas: Son importantes porque conectan la inmunidad innata con la adaptativa, presentando partes de los gérmenes a otras células.
- Mastocitos, eosinófilos y basófilos: Participan en la respuesta inflamatoria y en la defensa contra parásitos.
- Células asesinas naturales (NK): Atacan y destruyen células que están infectadas por virus o que se han vuelto anormales, como las células de un tumor.
Inmunidad Adaptativa o Adquirida
Este sistema es más avanzado y permite una respuesta más específica y con "memoria".
Linfocitos: Los Especialistas
Las células clave de la inmunidad adaptativa son los linfocitos, que se dividen en:
- Células B: Producen anticuerpos, que son como "llaves" que encajan con partes específicas de los gérmenes (antígenos) para marcarlos y neutralizarlos.
- Células T: Hay varios tipos:
* Linfocitos T citotóxicos: Matan células infectadas por virus o células anormales. * Linfocitos T colaboradores: No matan directamente, pero dirigen a otras células inmunitarias para que hagan su trabajo. Son como los "directores de orquesta" del sistema inmunitario. * Células T γ δ: Son un tipo especial que se encuentra entre la inmunidad innata y la adaptativa, reconociendo ciertos gérmenes sin necesidad de un procesamiento especial.

Memoria Inmunitaria
Cuando las células B y T se activan y se multiplican, algunas se convierten en "células de memoria" que viven mucho tiempo. Estas células recuerdan a cada germen que han encontrado. Si el mismo germen vuelve a entrar al cuerpo, estas células de memoria actúan rápidamente para eliminarlo. Esta es la base de la vacunación.
- Inmunidad pasiva: Es cuando recibes anticuerpos de otra fuente, como un bebé que recibe anticuerpos de su madre durante el embarazo o a través de la leche materna. Es de corta duración.
- Inmunidad activa: Se adquiere después de una infección o mediante la vacunación. Tu propio cuerpo produce las células de memoria y los anticuerpos, y dura mucho tiempo.
La vacunación es una forma muy efectiva de proteger a las personas. Al introducir una parte inofensiva de un germen, el cuerpo aprende a reconocerlo y a crear defensas sin enfermarse. Además, si muchas personas se vacunan, se crea la "inmunidad de rebaño", que protege a quienes no pueden vacunarse, como bebés o personas con sistemas inmunitarios debilitados.
Problemas del Sistema Inmunitario
A veces, el sistema inmunitario puede fallar:
- Inmunodeficiencias: Ocurre cuando una o más partes del sistema inmunitario no funcionan bien. Esto hace que el cuerpo sea más vulnerable a las infecciones. Puede ser heredada o adquirida, por ejemplo, por una nutrición deficiente o ciertas enfermedades.
- Autoinmunidad: El sistema inmunitario se confunde y ataca partes del propio cuerpo, como si fueran extrañas. Esto puede causar enfermedades como la artritis reumatoide o la diabetes mellitus tipo 1.
- Hipersensibilidad (Alergias): Es una respuesta inmunitaria exagerada que daña los propios tejidos del cuerpo. Las alergias son un ejemplo común, donde el cuerpo reacciona fuertemente a sustancias inofensivas como el polen.
Manipulación del Sistema Inmunitario en Medicina
Los científicos y médicos pueden influir en el sistema inmunitario para tratar enfermedades:
- Inmunosupresores: Son medicamentos que reducen la actividad del sistema inmunitario. Se usan para controlar enfermedades autoinmunes, inflamaciones graves o para evitar que el cuerpo rechace un órgano trasplantado.
- Estimulación: También se busca estimular el sistema inmunitario para que combata mejor ciertas enfermedades, como algunos tipos de cáncer. La inmunoterapia es un campo de investigación activo en este sentido.

Cómo los Gérmenes Engañan al Sistema Inmunitario
Los gérmenes han desarrollado trucos para evitar ser detectados y destruidos por el sistema inmunitario:
- Algunas bacterias secretan enzimas para romper las barreras del cuerpo o inyectan proteínas para desarmar las defensas.
- Muchos gérmenes, como los virus, se esconden dentro de las células del cuerpo, donde están protegidos de los ataques directos.
- Otros gérmenes cambian rápidamente sus características superficiales para que el sistema inmunitario no los reconozca, o se cubren con proteínas del propio cuerpo para "camuflarse".
Historia de la Inmunología
La inmunología es la ciencia que estudia el sistema inmunitario. Sus orígenes se remontan a la antigua Grecia, cuando se observó que las personas que se recuperaban de una enfermedad no volvían a enfermarse de la misma.
- En el siglo XIX, Louis Pasteur desarrolló las primeras vacunas, basándose en la idea de que los microbios causan enfermedades.
- Más tarde, científicos como Paul Ehrlich y Elie Metchnikoff hicieron grandes descubrimientos sobre cómo funcionan los anticuerpos y las células de defensa.
- En el siglo XX, se descubrieron más detalles sobre los diferentes tipos de células inmunitarias, cómo se comunican entre sí (a través de sustancias llamadas citocinas) y cómo el sistema inmunitario "recuerda" a los gérmenes.
Estos descubrimientos han sido fundamentales para entender cómo nos defendemos de las enfermedades y para desarrollar nuevas formas de tratamiento y prevención.
Galería de imágenes
-

Una imagen al microscopio electrónico de barrido de sangre humana normal circulante. Se pueden ver glóbulos rojos, varios glóbulos blancos incluyendo linfocitos, un monocito, un neutrófilo y muchas plaquetas pequeñas en forma de disco.
-
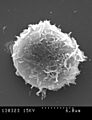
Los monocitos muestran una intensa actividad en su superficie celular.
-
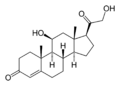
La corticosterona es un medicamento que puede afectar el sistema inmunitario.



Véase también
 En inglés: Immune system Facts for Kids
En inglés: Immune system Facts for Kids
