
Neutrófilo para niños
Datos para niños Neutrófilo |
||
|---|---|---|
 Imagen tomada con un microscopio óptico en la que se observa un neutrófilo rodeado de glóbulos rojos en un frotis sanguíneo. Tinción de May Grünwald-Giemsa.
|
||
 Representación 3D de un neutrófilo.
|
||
| Nombre y clasificación | ||
| Sinónimos |
|
|
| Latín | Granulocytus neutrophilus; Neutrophilus; Granulocytus neutrophilus segmentonuclearis | |
| TH | H2.00.04.1.02012 | |
| TH | H2.00.04.1.02012 | |
| Información anatómica | ||
| Sistema | Inmune | |
|
|
||
Los neutrófilos son un tipo de glóbulos blancos muy importantes para tu salud. También se les llama polimorfonucleares (PMN). Son las células blancas más comunes en la sangre humana, formando entre el 50% y el 60% de ellas. Su tamaño es muy pequeño, miden entre 9 y 12 micrómetros (μm).
La función principal de los neutrófilos es proteger tu cuerpo. Actúan como "soldados" que se comen y destruyen bacterias y hongos que pueden causar enfermedades. Este proceso se llama fagocitosis. Los neutrófilos son muy rápidos y pueden moverse fácilmente por el cuerpo.
Contenido
¿Cómo son los neutrófilos?
Los neutrófilos tienen características especiales que los hacen únicos.
Apariencia al microscopio
Cuando se observan bajo un microscopio óptico, los neutrófilos tienen un color rosa suave. Esto se debe a que no se tiñen con los colorantes ácidos ni básicos que se usan en el laboratorio.
Su núcleo celular (el "cerebro" de la célula) tiene una forma muy particular. Parece que está dividido en varias partes, de 2 a 5 lóbulos, conectados por pequeños puentes. En los neutrófilos más jóvenes, el núcleo se ve como una banda sin dividir.
El citoplasma (la parte gelatinosa que rodea el núcleo) contiene muchos gránulos pequeños de color púrpura. Estos gránulos guardan enzimas especiales y una sustancia llamada fagocitina. Todo esto es necesario para que el neutrófilo pueda luchar contra los gérmenes.
Producción y vida útil
Tu médula ósea (un tejido dentro de tus huesos) es como una fábrica de neutrófilos. En un adulto sano, se producen alrededor de 1200 millones de neutrófilos cada día. Sin embargo, su vida es corta, durando solo unas horas o pocos días.
¿Cómo defienden los neutrófilos tu cuerpo?
Los neutrófilos son muy activos y se mueven para protegerte.
Movimiento hacia la infección: Diapédesis
Los neutrófilos son células muy móviles. Su consistencia gelatinosa les permite atravesar las paredes de los vasos sanguíneos. Así pueden llegar a los tejidos donde hay una infección o inflamación. Este movimiento se llama diapédesis.
Cuando hay una infección, especialmente por bacterias, los neutrófilos son los primeros en llegar al lugar. Son guiados por señales químicas, como la interleucina-8 (IL-8) y el interferón-gamma (IFN-γ). Este proceso de guía se llama quimiotaxis. Por eso, los neutrófilos son las células más abundantes en el pus, que es una señal de que tu cuerpo está combatiendo una infección.
La liberación de neutrófilos desde los vasos sanguíneos es controlada por sustancias como la histamina y el TNF-α. Estas sustancias activan las células que recubren los vasos sanguíneos (llamadas endotelio), permitiendo que los neutrófilos se peguen y luego pasen a través de ellas. Pueden llegar a los tejidos infectados en tan solo 5 horas.
Reconocimiento de amenazas
Una vez que los neutrófilos llegan al lugar de la infección, necesitan saber qué atacar. Tienen "sensores" especiales en su superficie que les permiten reconocer a los microbios y las células dañadas.
- Receptores para microbios: Algunos receptores, como los receptores de tipo Toll (TLR), identifican partes específicas de bacterias y virus.
- Receptores para señales químicas: Otros receptores detectan péptidos (pequeñas proteínas) de bacterias o señales de daño celular.
- Receptores para opsoninas: Las opsoninas son proteínas de defensa que cubren a los microbios, marcándolos para que los neutrófilos los reconozcan y los eliminen más fácilmente. Los anticuerpos son un tipo de opsonina.
- Receptores para citoquinas: También tienen receptores para citoquinas, que son mensajes químicos que otras células del sistema inmune envían para activar a los neutrófilos.
Eliminación del agente dañino: Fagocitosis
Cuando un neutrófilo reconoce una amenaza, se activa para eliminarla. La fagocitosis es el proceso principal y tiene tres pasos:
1. Reconocimiento y unión: El neutrófilo se une al microbio usando sus receptores especiales. 2. Incorporación: El neutrófilo extiende partes de su cuerpo (como "brazos" llamados pseudópodo) para rodear al microbio. Luego, lo "traga" y lo encierra en una burbuja dentro de sí mismo, llamada fagosoma. Este fagosoma se une con otra burbuja llena de enzimas, el lisosoma, formando un fagolisosoma. 3. Destrucción: Dentro del fagolisosoma, el neutrófilo libera enzimas y sustancias químicas muy potentes. Estas sustancias, como el peróxido de hidrógeno (similar al agua oxigenada) y el hipoclorito (el ingrediente activo de la lejía), atacan y destruyen al microbio. Este proceso es muy efectivo para eliminar las amenazas.
¿Qué pasa después de la batalla?
Los neutrófilos tienen una vida corta. Después de cumplir su misión de destruir microorganismos, mueren por un proceso llamado apoptosis. La acumulación de neutrófilos muertos, bacterias muertas y líquido forma lo que conocemos como pus. Esto es una señal de que tu cuerpo está limpiando la infección.
A veces, en ciertas situaciones, los neutrófilos pueden dañar tejidos sanos si la respuesta inflamatoria es muy fuerte o se dirige por error contra el propio cuerpo. Sin embargo, su función principal es siempre protegerte de las infecciones.
Galería de imágenes
-
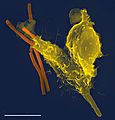
Imagen tomada con un microscopio electrónico en la que se observa un neutrófilo (amarillo) fagocitando una bacteria de carbunco (naranja).
-
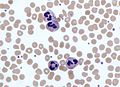
Imagen tomada con un microscopio óptico en la que se observan polimorfonucleares neutrófilos rodeados de glóbulos rojos en un frotis sanguíneo.
-
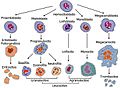
Linaje de las células sanguíneas.
-
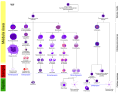
Linaje más completo.



_diagram_es.svg)
Véase también
 En inglés: Neutrophil Facts for Kids
En inglés: Neutrophil Facts for Kids
