
Apicomplexa para niños
Datos para niños Apicomplexa |
||
|---|---|---|
 Esporozoito de Plasmodium
|
||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) | Supergrupo SAR | |
| Superfilo: | Alveolata | |
| Filo: | Miozoa | |
| Subfilo: | Myzozoa | |
| Infrafilo: | Apicomplexa Levine, 1970 |
|
| Superclase: | Sporozoa Leuckart, 1879 |
|
| Subgrupos | ||
|
||
Los apicomplejos (Apicomplexa) son un gran grupo de protistas. Son organismos muy pequeños que viven dentro de las células de los animales. Se conocen unas 5000 especies diferentes.
Estos organismos tienen una característica especial llamada complejo apical. Es como una herramienta que les ayuda a entrar en las células de otros seres vivos. Son unicelulares, lo que significa que están hechos de una sola célula. También forman esporas, que son como semillas que les ayudan a sobrevivir y dispersarse.
Los apicomplejos no tienen estructuras para moverse, como colas (flagelos) o pseudópodos (extensiones de la célula), excepto en algunas etapas de sus gametos (células reproductivas). Este grupo incluye organismos como las gregarinas, coccidios y hemosporidios (como el Plasmodium). Algunos apicomplejos pueden causar enfermedades en animales y humanos, como la coccidiosis, la malaria y la babesiosis.
Contenido
- ¿Qué enfermedades causan los apicomplejos?
- ¿Cómo son los apicomplejos?
- ¿Cómo es el ciclo de vida de los apicomplejos?
- ¿Cómo se clasifican los apicomplejos?
- ¿Cómo se relacionan los apicomplejos con otros grupos?
- ¿Cómo evolucionaron los apicomplejos?
- ¿Por qué son importantes los apicomplejos en la medicina?
- Galería de imágenes
- Véase también
¿Qué enfermedades causan los apicomplejos?
Casi todos los apicomplejos son endoparásitos, lo que significa que viven dentro de los animales. Solo una especie, Nephromyces, vive en simbiosis con animales marinos, lo que significa que se ayudan mutuamente.
Algunas de las enfermedades que pueden causar en los seres humanos son:
- Coccidiosis, que incluye:
- Criptosporidiosis (causada por Cryptosporidium parvum)
- Ciclosporosis (causada por Cyclospora cayetanensis)
- Toxoplasmosis (causada por Toxoplasma gondii)
- Isosporiasis (causada por Cystoisospora belli)
- Malaria (causada por Plasmodium)
- Babesiosis (causada por Babesia)
¿Cómo son los apicomplejos?

A. Complejo Apical
1. Anillo polar
2. Intraconoide microtúbulo
3. Conoide
4. Anillo posterior
5. Complejo de membrana interna
6. Subpellicular microtúbulo
7. Roptrias, retienen las enzimas liberadas durante la penetración en el huésped
8. Micronemas, importante para la invasión de la célula huésped y la movilidad de deslizamiento
9. Mitocondria
10. Microporo
11. Gránulos densos
12. Membranas de apicoplasto, (4, rojo secundario, no fotosintético)
13. Aparato de Golgi
14. Núcleo
15. Retículo endoplasmático
Todos los apicomplejos tienen una etapa en su vida que puede causar una infección. En esta etapa, llamada esporozoito, poseen el complejo apical. Este complejo es un orgánulo especial que se encuentra en un extremo de la célula.
El complejo apical tiene tres partes principales:
- Un grupo de microtúbulos en espiral, llamado conoide.
- Unas estructuras que liberan sustancias, llamadas roptrias.
- Una o más bandas de microtúbulos, llamadas anillo polar.
También puede tener otras bolsitas secretoras llamadas micronemas. El complejo apical ayuda al organismo a entrar en la célula de su hospedador (el animal donde vive). Lo hace perforando la célula con el conoide y liberando sustancias químicas de las roptrias.
Además del complejo apical, tienen otros orgánulos esféricos llamados gránulos densos. Estos actúan después de que el parásito entra en la célula. La membrana de la célula del apicomplejo está reforzada por unas bolsitas llamadas alvéolos corticales, que le dan una forma semirrígida. Solo hay una pequeña abertura llamada microporo por donde se alimentan.
Los apicomplejos solo tienen flagelos (colas para moverse) en sus gametos (células reproductivas). Estos flagelos están en la parte trasera y pueden ser uno, dos o tres. Tienen una forma única de moverse, deslizándose, lo que les permite atravesar tejidos y entrar y salir de las células. Esto lo logran usando unas estructuras que se adhieren y unos pequeños motores internos.
La mayoría de los apicomplejos tienen un apicoplasto. Este es un tipo de plástido (un orgánulo celular) que no realiza la fotosíntesis. Otras características comunes de este grupo son la reproducción sexual, el uso del microporo para alimentarse y la producción de ooquistes que contienen esporozoitos.
La célula de los apicomplejos tiene un núcleo con una sola copia de sus genes (haploide). También tiene un aparato de Golgi y mitocondrias con crestas tubulares. No tienen centriolos ni cloroplastos. La célula está cubierta por una capa de tres membranas. Su mitosis (división celular) es especial, y la meiosis (división para la reproducción sexual) ocurre en el cigoto.
¿Cómo es el ciclo de vida de los apicomplejos?


La mayoría de los apicomplejos tienen un ciclo de vida complicado. Incluye tanto la reproducción asexual como la sexual.
Generalmente, un huésped (el animal donde vive el parásito) se infecta al comer quistes. Dentro del huésped, estos quistes se dividen y producen esporozoitos. Los esporozoitos entran en las células del huésped.
Con el tiempo, las células infectadas se rompen, liberando merozoitos. Estos merozoitos infectan nuevas células. Este proceso puede repetirse varias veces. Finalmente, se forman gametocitos, que son células que darán lugar a los gametos. Los gametos se unen para formar nuevos quistes, completando el ciclo.
Existen muchas variaciones de este ciclo básico. Muchos apicomplejos necesitan más de un huésped para completar su ciclo de vida.
¿Cómo se clasifican los apicomplejos?
Los apicomplejos se dividen en cuatro grupos principales: gregarinas, coccidios, hemosporidios y piroplasmas. Las gregarinas y los coccidios están relacionados (forman la clase Conoidasida). Los hemosporidios y los piroplasmas también están relacionados (forman la clase Aconoidasida).
Gregarinas
Las gregarinas suelen ser parásitos de gusanos anillados (anélidos), artrópodos (como insectos) y moluscos. A menudo se encuentran en el intestino de sus huéspedes, pero pueden vivir en otros tejidos.
En su ciclo de vida, una forma llamada trofozoito crece dentro de la célula del huésped. Luego se divide muchas veces para formar merozoitos. Estos merozoitos salen de la célula y pueden infectar otras. En algún momento, se forman gametocitos. Estos gametocitos producen gametos que se unen para formar ooquistes. Los ooquistes salen del huésped para buscar uno nuevo.
Coccidios
Los coccidios son generalmente parásitos de animales con columna vertebral (vertebrados). Al igual que las gregarinas, suelen vivir en las células del intestino, pero también pueden infectar otros tejidos. Algunos son parásitos de la sangre.
El ciclo de vida de los coccidios es similar al de las gregarinas. La diferencia principal es cómo se forma el cigoto. Algunos trofozoítos crecen y se convierten en macrogametos (células reproductoras grandes), mientras que otros se dividen para formar microgametos (células reproductoras pequeñas y móviles). Los microgametos buscan y fertilizan a los macrogametos. El macrogameto fertilizado se convierte en un cigoto que forma un ooquiste, el cual suele salir del huésped.
Hemosporidios
Los hemosporidios tienen ciclos de vida más complejos. Alternan entre un huésped artrópodo (como un insecto) y un huésped vertebrado. Los trofozoitos viven en los eritrocitos (glóbulos rojos) u otros tejidos del huésped vertebrado.
Los gametos de los hemosporidios siempre se encuentran en la sangre. Cuando un insecto pica al vertebrado y se alimenta de su sangre, absorbe estos gametos. Dentro del insecto, los gametos se unen y forman un oocineto. Este oocineto se mueve y penetra en el cuerpo del insecto. Luego se transforma en un ooquiste y se divide para formar esporozoitos. Los esporozoitos salen del ooquiste y viajan a las glándulas salivales del insecto. Desde allí, son inyectados en un nuevo huésped vertebrado cuando el insecto lo pica.
Piroplasmas
Los piroplasmas tienen ciclos de vida muy parecidos a los hemosporidios. La diferencia es que los piroplasmas no forman ooquistes ni esporas. Además, en la etapa de trofozoito, están separados del eritrocito por una sola membrana, a diferencia de otros parásitos de la sangre que suelen tener al menos dos.
¿Cómo se relacionan los apicomplejos con otros grupos?
Los apicomplejos pertenecen a un grupo llamado Alveolata debido a la presencia de alvéolos corticales y otras características. Algunos organismos con flagelos, como Perkinsus y Colpodella, tienen estructuras parecidas al complejo apical. Antes se incluían en este grupo, pero ahora se cree que están más relacionados con los Dinoflagellata. Estos organismos podrían ser similares a los ancestros comunes de ambos grupos.
Otra conexión con los dinoflagelados es la presencia de un plasto especial en las células de los apicomplejos, llamado apicoplasto. Este orgánulo está rodeado por tres o cuatro membranas y se cree que es importante para la supervivencia del organismo, ayudando en la producción de lípidos. Se piensa que los apicoplastos tienen un origen común con los cloroplastos de los dinoflagelados.
Los apicomplejos forman parte de un grupo de organismos que antes se llamaban esporozoos. Los esporozoos eran protozoos parásitos que no tenían flagelos, pseudópodos ni cilios. Otros grupos que antes se clasificaban como esporozoos ahora se encuentran en diferentes categorías, como Ascetosporea, Myxozoa y Microsporidia. A veces, el término esporozoo se usa como sinónimo de Apicomplexa.
¿Cómo evolucionaron los apicomplejos?

El ciclo de vida complejo de los apicomplejos se entiende mejor al estudiar su evolución. Los estudios sugieren que los Apicomplexa se originaron de los Dinoflagellata, que son un gran grupo de protozoos que realizan la fotosíntesis.
Es probable que los ancestros de los apicomplejos fueran organismos que producían su propio alimento. Luego, desarrollaron la capacidad de entrar en las células intestinales de los animales que los comían. Con el tiempo, perdieron la capacidad de hacer fotosíntesis. Algunas especies de dinoflagelados actuales pueden vivir dentro de medusas y seguir haciendo fotosíntesis, porque los órganos de las medusas son casi transparentes. Pero en otros animales con órganos opacos, la capacidad de fotosíntesis se habría perdido rápidamente.
Se cree que los apicomplejos comenzaron infectando la pared intestinal de sus huéspedes, transmitiéndose a través de las heces. En algún momento, el parásito evolucionó y pudo infectar el hígado del huésped. Este patrón se ve en el género Cryptosporidium.
Más tarde, un apicomplejo desarrolló la capacidad de infectar las células de la sangre y de sobrevivir en los mosquitos. Una vez que la transmisión por mosquitos se estableció, la forma anterior de transmisión por heces se perdió. La teoría actual (2007) sugiere que géneros como Plasmodium evolucionaron de especies de Leukocytozoon. Los parásitos del género Leukocytozoon infectan los leucocitos (glóbulos blancos) y las células del hígado y el bazo. Se transmiten por medio de moscas negras (Simulium), que son un tipo de mosca relacionada con los mosquitos.
Los glóbulos blancos, la mayoría de las células del hígado y las del bazo pueden "comer" partículas (proceso llamado fagocitosis), lo que facilita la entrada del parásito. Sin embargo, cómo entran los parásitos del género Plasmodium en los eritrocitos (glóbulos rojos) aún no está claro, ya que lo hacen en menos de 30 segundos. No se sabe si este mecanismo evolucionó antes de que los mosquitos se convirtieran en los principales transmisores de Plasmodium.
El patrón de alternar entre reproducción sexual y asexual es muy común en los parásitos. Las ventajas de este tipo de ciclo de vida fueron reconocidas por Mendel. En condiciones favorables, la reproducción asexual es mejor porque los descendientes son muy parecidos a sus padres y están bien adaptados al ambiente. Pero en situaciones de estrés, como cuando el parásito se transfiere a un nuevo huésped, la reproducción sexual es mejor. Esto se debe a que mezcla los genes, lo que puede producir individuos mejor adaptados al nuevo entorno.
¿Por qué son importantes los apicomplejos en la medicina?
Los Apicomplexa incluyen muchos organismos que causan enfermedades importantes en humanos y animales domésticos. A diferencia de las bacterias que causan enfermedades, estos parásitos son eucariontes. Esto significa que sus células son más complejas y comparten muchas características con las células de sus huéspedes. Por esta razón, es muy difícil encontrar tratamientos que dañen al parásito sin dañar también al huésped.
Actualmente, no hay vacunas o tratamientos completamente efectivos para la mayoría de las enfermedades causadas por estos parásitos. Un posible objetivo para los medicamentos es el plástido que tienen. De hecho, algunos medicamentos que ya existen y son efectivos contra los apicomplejos, como las tetraciclinas, parecen actuar sobre este plástido.
La investigación sobre estos parásitos es complicada. A veces es difícil mantenerlos vivos en el laboratorio y estudiarlos. Sin embargo, recientemente se ha logrado estudiar el genoma (todo el material genético) de varias especies. Esto abre nuevas oportunidades para entender mejor cómo evolucionaron y cómo funcionan estos parásitos.
Galería de imágenes
-

Lankesteria (Gregarinasina)
-

Eugregarina septada (Gregarinasina)
-
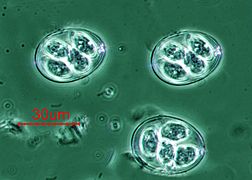
Quistes de Eimeria maxima (Coccidiasina)
-
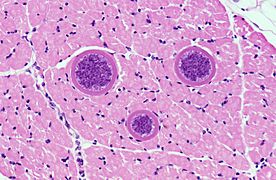
Quistes de Sarcocystis hominis (Coccidiasina)
-
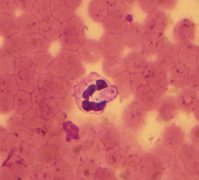
Gamonte de Hepatozoon canis (Coccidiasina)
-
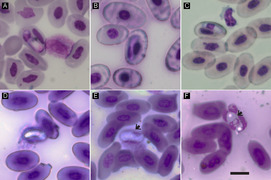
Gamontes de Hepatozoon domerguei (Coccidiasina)
-
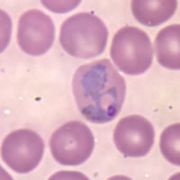
Trofozoito de Plasmodium dentro de un eritrocito (Haemosporida)
-
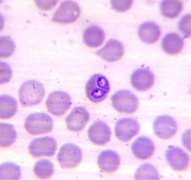
Dos trofozoitos de Babesia vogeli dentro de un eritrocito (Piroplasmida)
-
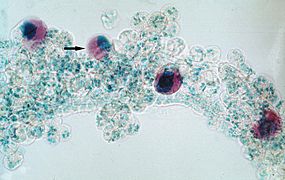
Esporoblasto de Theileria annulata (Piroplasmida)









Véase también
 En inglés: Apicomplexa Facts for Kids
En inglés: Apicomplexa Facts for Kids
