
Megalosaurus bucklandii para niños
Datos para niños Megalosaurus bucklandii |
||
|---|---|---|
| Rango temporal: 166 Ma Jurásico Medio | ||
 |
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Archosauria | |
| (sin rango): | Tetanurae | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Superfamilia: | Megalosauroidea | |
| Familia: | Megalosauridae | |
| Subfamilia: | Megalosaurinae | |
| Género: | Megalosaurus Buckland, 1824 |
|
| Especie: | M. bucklandii Mantell, 1827 |
|
| Sinonimia | ||
|
||
El Megalosaurus bucklandii es un dinosaurio terópodo extinto. Su nombre significa "lagarto grande" en latín y griego. Vivió hace unos 166 millones de años, durante el Jurásico Medio, en lo que hoy es Europa.
Fue el primer género de dinosaurio en ser descrito científicamente. William Buckland le dio el nombre Megalosaurus en 1824, por su gran tamaño. La especie, M. bucklandii, fue nombrada por Gideon Mantell en 1827 en honor a Buckland. Este dinosaurio está relacionado con otros como el Torvosaurus de América del Norte y el Poekilopleuron de Francia.
Durante un tiempo, muchos dinosaurios carnívoros grandes fueron clasificados erróneamente como Megalosaurus. Sin embargo, en años recientes, los científicos han estudiado mejor este género. Ahora, la mayoría de esas especies han sido reclasificadas en otros grupos.
Contenido
¿Cómo era el Megalosaurus?
El Megalosaurus era un depredador que caminaba sobre dos patas (bípedo). Medía unos 9 metros de largo y pesaba alrededor de una tonelada. Tenía un cuerpo equilibrado, ideal para correr. Sus vértebras del cuello le daban mucha flexibilidad.
Como todos los terópodos, tenía tres dedos hacia adelante en sus patas traseras. Sus patas delanteras eran pequeñas, pero fuertes, y probablemente tenían tres dedos.
Apariencia general del Megalosaurus
El Megalosaurus tenía la forma típica de un gran terópodo. Su cuerpo era horizontal y su larga cola le ayudaba a mantener el equilibrio. Sus patas traseras eran largas y fuertes. Las patas delanteras eran más cortas, pero muy robustas.
Al ser un carnívoro, tenía una cabeza grande y alargada. Sus dientes eran largos y afilados, como dagas, perfectos para cortar la carne de sus presas. Su esqueleto era muy fuerte y musculoso.
¿Qué tamaño tenía el Megalosaurus?

Tradicionalmente, se estimaba que el Megalosaurus medía unos 9 metros de largo. Sin embargo, como no se ha encontrado un esqueleto completo, es difícil saber su tamaño exacto.
Algunos científicos, como David Bruce Norman en 1984, pensaron que medía entre 7 y 8 metros. Gregory S. Paul estimó su peso en 1,1 toneladas en 1988. Investigaciones más recientes, como la de Paul en 2010, sugieren que podría haber medido 6 metros de largo y pesado 700 kilogramos.
Sin embargo, Roger Benson afirmó en 2010 que, aunque de tamaño mediano, el Megalosaurus seguía siendo uno de los terópodos jurásicos más grandes. Un hueso del muslo de 803 milímetros de largo sugiere un peso de 943 kilogramos. Otro hueso del muslo, de unos 86 centímetros, indica que algunos individuos eran aún más grandes.
Descubrimiento e investigación del Megalosaurus
El Megalosaurus es el primer dinosaurio nombrado en la historia moderna. Antes de la clasificación científica actual, algunos fósiles que hoy se consideran de Megalosaurus ya habían sido mencionados.
Solo la especie original, Megalosaurus bucklandii, se considera válida. Sus fósiles se han encontrado en las formaciones Corallian Oolite y Taynton Limestone del Jurásico Medio en Oxfordshire y Gloucestershire, al sudeste de Inglaterra.
Los primeros hallazgos de huesos
El primer fragmento de hueso de este género se encontró en una cantera de caliza en Cornwell, Inglaterra, en 1677. Este fragmento fue enviado a Robert Plot, un profesor de la Universidad de Oxford. Plot publicó una descripción en su libro Historia Natural de Oxfordshire en 1676.
Plot identificó correctamente el hueso como la parte inferior del fémur de un animal grande. Se dio cuenta de que era demasiado grande para cualquier especie conocida. Al principio, pensó que era el hueso del muslo de un elefante de guerra romano o de un humano gigante. Aunque el hueso se perdió, la ilustración de Plot es lo suficientemente detallada como para identificarlo como un fémur de Megalosaurus. Fue la primera ilustración publicada de un hueso de dinosaurio.
En 1763, el grabado de Plot se usó en un libro de Richard Brookes. Brookes le dio un nombre inusual, "... humanum", debido a su forma. Este nombre ya no se usa.
La investigación de William Buckland

A principios del siglo XIX, se hicieron más descubrimientos en la cantera de Stonesfield, cerca de Oxford. Estos huesos fueron adquiridos por el profesor William Buckland, de la Universidad de Oxford.
En 1818, el anatomista francés Georges Cuvier visitó a Buckland y confirmó que estos huesos pertenecían a un reptil gigante. Buckland publicó su descripción en 1824, afirmando que era un reptil gigante que caminaba sobre dos patas. Lo nombró Megalosaurus.

Buckland estudió los restos con su amigo William Conybeare. El 20 de febrero de 1824, Buckland anunció formalmente el nombre Megalosaurus. Fue el primer género de dinosaurio no aviar en ser nombrado.
En 1827, Gideon Mantell le dio el nombre completo de la especie: Megalosaurus bucklandii. Sin embargo, no fue hasta 1842 que Richard Owen acuñó el término "Dinosauria" para agrupar a estas criaturas.
Al principio, Buckland pensó que el Megalosaurus era un animal de cuatro patas que podía nadar y caminar en tierra. Lo imaginó como un lagarto gigantesco. Estimó que medía unos 12 metros de largo y pesaba como un elefante.
Reconstrucciones antiguas

En 1852, Benjamin Waterhouse Hawkins construyó un modelo de Megalosaurus de tamaño real para una exposición en el Crystal Palace Park en Sydenham, Londres. Esta estatua, que aún se conserva, mostraba al Megalosaurus como un animal de cuatro patas, similar a un mamífero.
La idea de que los dinosaurios carnívoros eran cuadrúpedos fue cuestionada más tarde. En 1870, se descubrió un esqueleto bastante completo de un gran terópodo, el Eustreptospondylus, que claramente caminaba sobre dos patas. Esto ayudó a los científicos a entender mejor la postura del Megalosaurus.
El Museo de Historia Natural de la Universidad de Oxford tiene la mayoría de los fósiles originales descritos por Buckland.
Clasificación del Megalosaurus
En 1824, Buckland clasificó al Megalosaurus dentro de los Sauria, pensando que estaba relacionado con los lagartos modernos. En 1842, Owen lo incluyó en el nuevo grupo de los Dinosauria.
En 1850, se creó una familia separada, los Megalosauridae, con Megalosaurus como el género principal. A lo largo de los años, su clasificación ha cambiado. Ahora se le considera parte de la superfamilia Megalosauroidea y la familia Megalosauridae.
Los científicos usan un método llamado cladística para entender cómo se relacionan los dinosaurios. En 2012, Matthew Carrano y sus colegas mostraron que Megalosaurus estaba estrechamente relacionado con Torvosaurus dentro de la subfamilia Megalosaurinae.
¿Dónde vivía el Megalosaurus?
El Megalosaurus vivió en lo que hoy es Europa durante el Período Jurásico. Pudo haber cazado estegosáuridos y saurópodos.
Las antiguas descripciones que muestran al Megalosaurus cazando Iguanodon no son correctas. Los fósiles de Iguanodon se encuentran en rocas mucho más jóvenes, del Cretácico Inferior.
Los únicos fósiles confirmados de Megalosaurus bucklandii provienen de Inglaterra. Compartía su hábitat con otros terópodos como Cruxicheiros y Iliosuchus, y con grandes saurópodos como Cetiosaurus. También se han encontrado restos de pterosaurios (reptiles voladores) y otros dinosaurios en los mismos lugares.
Debido a su tamaño y a que era común en su hábitat, se cree que el Megalosaurus era el principal depredador de su ecosistema.
El Megalosaurus en la cultura popular
El Megalosaurus fue el primer dinosaurio en aparecer en la cultura popular.
- En la novela de Charles Dickens, Casa desolada, el Megalosaurus es mencionado al principio para describir una niebla muy antigua.
- También aparece en la novela de John Brosnan de 1984, Carnosaurio.
- En la serie japonesa Dino Rey, un Megalosaurus es uno de los dinosaurios creados por el Dr. Z.
- La serie de televisión Dinosaurios (1991-1994) presenta a Earl Sinclair, un Megalosaurus, como el padre de una familia de dinosaurios.
Galería de imágenes
-

Tamaño comparativo. Humano en azul, lectotipo en rosa, espécimen más grande en rojo.
-
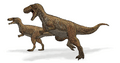
Reconstrucción de un Megalosaurus.
-

Grabado antiguo de dos Megalosaurus con la clásica postura (incorrecta) de la época.
-

Dinosaurios en lucha (1865).
-

Reconstrucción en el Crystal Palace, en Londres. Las primeras reconstrucciones lo muestran como un cuadrúpedo, pero las modernas lo representan bípedo.
,_plate_XXI_-_BL.jpg)
Véase también
 En inglés: Megalosaurus Facts for Kids
En inglés: Megalosaurus Facts for Kids
