
Theropoda para niños
Datos para niños Terópodos |
||
|---|---|---|
| Rango temporal: 231,4 Ma - 0 Ma Triásico Superior - Holoceno | ||
 Diversidad de terópodos.
|
||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: | Animalia | |
| Subreino: | Eumetazoa | |
| Superfilo: | Deuterostomia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Infrafilo: | Gnathostomata | |
| Superclase: | Tetrapoda | |
| Clase: | Sauropsida | |
| Subclase: | Eureptilia | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda Marsh, 1881 |
|
| Subgrupos | ||
|
||
Los terópodos (del griego therion = bestia y podos = pie, que significa «pie de bestia») son un grupo de dinosaurios. Se caracterizan por tener huesos huecos y patas con tres dedos que usaban para caminar.
Originalmente, se pensaba que todos los terópodos eran depredadores. Sin embargo, con el tiempo, algunos grupos evolucionaron para comer plantas (herbívoros), de todo (omnívoros), peces (piscívoros) o insectos (insectívoros).
Los terópodos aparecieron hace unos 231.4 millones de años, al final del período Triásico. Fueron los únicos grandes carnívoros terrestres desde el Jurásico Inferior hasta el final del Cretácico, hace unos 66 millones de años. Las aves que conocemos hoy en día, con sus cerca de 10.500 especies, evolucionaron de pequeños terópodos.
Contenido
¿Cómo vivían los terópodos?
¿Qué comían los terópodos?

La dieta de los terópodos era muy variada. Aunque al principio se creía que todos comían carne, los descubrimientos recientes muestran que muchos tenían dietas diferentes.
Los primeros fósiles de terópodos que se encontraron tenían dientes afilados, perfectos para cortar carne. Algunos incluso mostraban pruebas directas de lo que habían comido. Por ejemplo, se encontró un fósil de Compsognathus longipes con restos de un lagarto en su estómago. También se halló un Velociraptor mongoliensis en plena lucha con un Protoceratops.
Los primeros terópodos que no comían carne fueron los tericinosaurios. Estos dinosaurios tenían grandes barrigas para digerir plantas y cabezas pequeñas con pico y dientes en forma de hoja. Otros grupos de terópodos también cambiaron su dieta, comiendo semillas (como algunos trodóntidos) o insectos (como muchos avialanos y alvarezsaurios).
Algunos terópodos, como los ovirraptorosaurios y ornitomimosaurios, eran posiblemente omnívoros. Otros, como Masiakasaurus y los espinosáuridos, se especializaron en comer peces.
Los científicos descubren la dieta de los terópodos estudiando sus dientes, las marcas que dejaban en los huesos de sus presas y lo que se encuentra en sus estómagos fosilizados. Algunos terópodos, como Baryonyx, usaban gastrolitos (piedras en el estómago) para ayudar a moler su comida.
La mayoría de los dientes de terópodos eran como cuchillos, con bordes aserrados. Esto les ayudaba a cortar la carne. Investigaciones recientes han mostrado que unas "grietas" en sus dientes eran en realidad pliegues que los hacían más fuertes y evitaban que se rompieran al morder.
¿Cómo era su piel y sus plumas?

Los terópodos tenían una piel muy variada. Se ha confirmado que muchos tenían plumas o estructuras parecidas a plumas. Sin embargo, en los terópodos grandes, las plumas solían estar solo en los jóvenes o en ciertas partes del cuerpo.
Muchos terópodos grandes tenían la piel cubierta de pequeñas escamas. Algunas especies tenían también escamas más grandes con huesos dentro, llamadas osteodermos. Un ejemplo es el Carnotaurus, del que se han encontrado impresiones detalladas de su piel.
Los terópodos más cercanos a las aves tenían plumas cortas y simples. Otros, como los tericinosaurios, tenían plumas grandes y rígidas. Los terópodos con más plumas, como los dromeosáuridos, solo tenían escamas en los pies. Algunas especies podían tener una mezcla de escamas y plumas. Por ejemplo, Scansoriopteryx tenía escamas en la cola, y Juravenator pudo haber tenido escamas con algunos filamentos de plumas. Otros, como el Anchiornis, estaban completamente cubiertos de plumas, incluso en los dedos de los pies.
¿Qué tan grandes eran los terópodos?

Durante mucho tiempo, el Tyrannosaurus fue considerado el terópodo más grande. Pero se han descubierto otros dinosaurios carnívoros gigantes, como el Spinosaurus, Carcharodontosaurus y Giganotosaurus.
Se cree que el Spinosaurus era más largo que el Tyrannosaurus, quizás unos 3 metros más. Sin embargo, el Tyrannosaurus pudo haber sido más alto y pesado. Los ejemplares de Tyrannosaurus llamados "Sue" y "Scotty" son los terópodos más pesados conocidos. No se sabe por qué estos animales crecieron tanto, superando a otros depredadores terrestres.
El terópodo más grande que vive hoy es el avestruz, que puede medir hasta 2.74 metros de altura y pesar entre 63.5 y 145.15 kilogramos.
El terópodo no aviano más pequeño conocido es el Anchiornis, que pesaba solo 110 gramos y medía 34 centímetros. Si incluimos las aves modernas, el colibrí zunzuncito es el más pequeño, con 1.9 gramos y 5.5 centímetros de largo.
Una teoría sugiere que el tamaño de los terópodos disminuyó continuamente durante 50 millones de años. Pasaron de un promedio de 163 kilogramos a solo 0.8 kilogramos en la línea que dio origen a las aves modernas. Sus esqueletos cambiaron cuatro veces más rápido que los de otros dinosaurios.
¿Cómo se movían los terópodos?
.jpg)
Todos los terópodos conocidos caminaban sobre dos patas (eran bípedos). Sus patas delanteras eran más cortas y se usaban para diferentes tareas. En las aves modernas, el cuerpo se mantiene bastante erguido, y la parte superior de la pata (fémur) está casi paralela a la columna vertebral.
En el pasado, se pensaba que terópodos como los carnosaurios y tiranosáuridos caminaban muy erguidos, casi como los canguros, usando sus colas como apoyo. Pero estudios más recientes, desde los años 70, sugieren que adoptaban una postura más horizontal, manteniendo la cola en el aire.
Aún hay debate sobre cómo se orientaban sus patas. Algunos creen que el fémur estaba vertical, mientras que otros piensan que la rodilla estaba muy doblada al caminar, incluso en los terópodos gigantes. Es probable que hubiera muchas formas diferentes de caminar y posturas en los distintos grupos de terópodos.
¿Cómo era su cerebro y sus sentidos?
Aunque son raros, se han encontrado fósiles de cerebros completos de terópodos. También se pueden reconstruir usando tecnología de tomografía axial computarizada (TAC) y programas 3D. Estos descubrimientos son muy importantes porque nos ayudan a entender cómo evolucionó el cerebro de las aves a partir de los reptiles antiguos. El cerebro de los terópodos se hizo más grande, especialmente en los Coelurosauria y luego en los maniraptoranos y las primeras aves.
¿Cómo eran sus patas delanteras?
_fig._1.png)
Las patas delanteras cortas en comparación con las traseras eran comunes en terópodos como los abelisáuridos (por ejemplo, Carnotaurus) y los tiranosáuridos (como Tyrannosaurus). Sin embargo, no todos eran así. Los espinosáuridos y muchos celurosaurios tenían patas delanteras bien desarrolladas.
Las manos de los terópodos también eran muy diversas. La mayoría de los terópodos no avianos tenían tres dedos con garras afiladas. Algunos terópodos más antiguos, como Herrerasaurus, tenían cuatro dedos. Los ceratosaurios solían tener cuatro dedos, mientras que muchos tetanuranos tenían tres.
Se cree que las patas delanteras se usaban de diferentes maneras. Los espinosáuridos podrían haber usado sus fuertes brazos para atrapar peces. Algunos pequeños maniraptoranos, como los escansoriopterígidos, podrían haber usado sus patas para trepar árboles. Las patas delanteras de la mayoría de las aves modernas se usan para volar, aunque algunas, como las de los pingüinos, están adaptadas para nadar.
¿Cómo movían sus patas delanteras?

A diferencia de cómo se muestran a menudo en películas, los terópodos tenían un movimiento muy limitado en sus patas delanteras. No podían girar sus antebrazos para que las palmas miraran hacia el suelo o hacia atrás, como lo hacemos los humanos. Esto se debe a que uno de los huesos del antebrazo (el radio) estaba bloqueado en el otro (el cúbito).
La muñeca también tenía poca movilidad en muchas especies, lo que hacía que el antebrazo y la mano se movieran como una sola pieza. La única forma en que la palma de un terópodo podía mirar hacia abajo era extendiendo los brazos hacia los lados, como hacen las aves al levantar sus alas.
En los carnosaurios como Acrocanthosaurus, la mano era bastante flexible, con dedos que podían moverse. Esto también ocurría en terópodos más antiguos. Sin embargo, en los celurosaurios, como los ornitomimosaurios y especialmente los dromeosáuridos, la mano perdió mucha flexibilidad y tenía dedos muy rígidos. Los dromeosáuridos y otros maniraptoranos tenían una muñeca más móvil gracias a un hueso especial en forma de media luna, que les permitía doblar la mano hacia atrás, como en las aves modernas.
¿Cómo se clasifican los terópodos?
Los terópodos son un grupo muy grande que incluye a todos los dinosaurios carnívoros del Triásico, Jurásico y Cretácico, y también a sus descendientes directos: las aves.
Los terópodos se dividen en dos grupos principales: los ceratosaurios y los tetanuros. También están los herrerasaurios, que son los más antiguos.
- Los ceratosaurios incluyen a los primeros terópodos grandes, como "Ceratosaurus" del Jurásico temprano, y a los abelisáuridos, que vivieron en el Cretácico en lugares como Sudamérica, África y la India.
- Los tetanuros son un grupo más evolucionado y diverso. Incluyen a los "carnosaurios" (grandes depredadores del Jurásico y Cretácico) y a los "celurosaurios", que van desde los ágiles dromeosáuridos hasta el famoso "Tyrannosaurus rex".
Galería de imágenes
-

Diversidad de terópodos.
-

Espécimen del trodóntido Jinfengopteryx elegans, con semillas preservadas en su región estomacal.
-

Fósil de Anchiornis, que muestra impresiones preservadas de grandes plumas.
-

Comparación de tamaño de algunos terópodos gigantes.
-
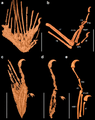
Ala momificada de un enantiornite preservada en ámbar del Cretácico.
-
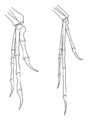
Diagrama de las extremidades delanteras de Deinonychus (izquierda) y Archaeopteryx (derecha) ilustrando la postura tipo ala.
Véase también
 En inglés: Theropod Facts for Kids
En inglés: Theropod Facts for Kids
