
Evolución humana para niños
La evolución humana es el fascinante viaje de nuestra especie, Homo sapiens, desde sus primeros ancestros hasta cómo somos hoy. Es como una gran historia que se cuenta a través de pistas encontradas en la genética, los fósiles, la arqueología y otras ciencias.
Cuando hablamos de "humanos" en esta historia, nos referimos a las especies del grupo Homo. Pero la evolución humana también incluye a otros parientes cercanos como los Ardipithecus y los Australopithecus.
Los científicos creen que el ancestro común de los humanos modernos y los chimpancés vivió en África hace entre cinco y siete millones de años. Desde entonces, muchas especies de homininos (nuestros parientes más cercanos) han aparecido y desaparecido, ¡excepto nosotros, Homo sapiens!
| Los hominoideos descendemos de un antepasado común |
|---|
| Antepasados de Homo sapiens |
| Anterior al género Homo |
|
Primeros Homínidos |
| Género Homo |
|
Contenido
¿Qué nos dice la genética sobre la evolución humana?
Al estudiar nuestro genoma (el conjunto de toda nuestra información genética), los científicos han descubierto cosas muy interesantes. Por ejemplo, los Homo sapiens compartimos casi el 99% de nuestros genes con los chimpancés y los bonobos. Esto muestra lo cerca que estamos de ellos en el árbol de la vida.
¿Hubo cruces entre especies humanas antiguas?
Los estudios genéticos sugieren que, en el pasado, hubo cruces entre diferentes especies de homínidos. Por ejemplo, las personas de origen no africano tienen entre un 1% y un 4% de genes de los Homo neanderthalensis (neandertales). También se ha encontrado ADN de los denisovanos en poblaciones actuales de lugares como Papúa Nueva Guinea. Estos genes antiguos podrían haber ayudado a los humanos a adaptarse a nuevos ambientes.
¿Qué son los retrovirus endógenos humanos?
Una parte importante de nuestro genoma (casi el 8%) está formada por "retrovirus endógenos humanos" (HERV). Son como restos de virus antiguos que se integraron en nuestro ADN a lo largo de la evolución. ¡Es como si nuestro cuerpo guardara un registro de encuentros con virus del pasado!
Etapas clave en la evolución humana
La historia de la evolución humana se divide en varias etapas, marcadas por la aparición de nuevas especies y cambios importantes.
Los primeros homínidos bípedos
Los primeros posibles homínidos que caminaban sobre dos patas (bípedos) aparecieron hace unos 7 millones de años en África. Algunos de ellos son Sahelanthropus tchadiensis, Orrorin tugenensis y Ardipithecus. Los fósiles de estos primeros homínidos son escasos. Sin embargo, el descubrimiento de un esqueleto casi completo de Ardipithecus ramidus, apodado "Ardi", mostró que podía caminar erguido, aunque sus pies aún tenían un dedo gordo que le ayudaba a agarrarse a las ramas.
Los Australopitecinos: Caminantes seguros
Los Australopithecus fueron los primeros homínidos de los que estamos seguros que caminaban completamente erguidos. Vivieron en las sabanas de África Oriental hace entre 4 y 2.5 millones de años. Tuvieron mucho éxito y se diversificaron en varias especies.
Cuando el clima cambió y las sabanas se volvieron más secas, algunos Australopithecus se especializaron en comer plantas duras, dando origen a los Paranthropus. Otros se hicieron más carnívoros, lo que llevó al surgimiento de las primeras especies del género Homo.
Los primeros Homo: Fabricantes de herramientas
No se sabe con exactitud de qué especie de Australopithecus surgieron los primeros Homo. Lo que sí sabemos es que los primeros Homo eran capaces de fabricar herramientas de piedra muy simples, llamadas "Olduvayense". Las herramientas más antiguas tienen unos 2.6 millones de años.
De esta etapa, conocemos dos especies principales: Homo rudolfensis y Homo habilis, que vivieron en África Oriental hace entre 2.5 y 1.8 millones de años. Su cerebro era más grande que el de los Australopitecinos.
¿Cómo se poblaron Europa y Asia?
La siguiente especie importante fue Homo ergaster, que apareció hace unos 1.8 millones de años. Se cree que fue el primero de nuestros ancestros en salir de África. Se han encontrado fósiles de Homo georgicus en Georgia (Asia), que son muy antiguos y muestran esta primera migración.
Esta migración llevó a la aparición de diferentes grupos: Homo erectus en Asia (China, Java) y Homo antecessor en Europa (España, Italia). Mientras tanto, los Homo ergaster que se quedaron en África desarrollaron herramientas más avanzadas, llamadas "Achelense".
En Europa, se han encontrado evidencias de presencia humana desde hace más de 1 millón de años. Los últimos homínidos de esta fase fueron Homo heidelbergensis en Europa (posible ancestro de los neandertales) y Homo rhodesiensis en África (posible ancestro del hombre moderno).
Nuevos orígenes en África: La llegada de Homo sapiens
La etapa final de nuestra evolución incluye a tres especies humanas inteligentes que convivieron: el Homo neanderthalensis (neandertal), el homínido de Denisova y el Homo sapiens (nosotros).
Los neandertales evolucionaron en Europa y Oriente Medio hace unos 230,000 años, adaptándose al clima frío. Los denisovanos vivieron en Asia hace unos 40,000 años.
Los fósiles más antiguos de Homo sapiens se han encontrado en Marruecos (hace 300,000 años) y Etiopía (hace unos 200,000 años). Esto sugiere que África es la "cuna de la humanidad".
Hace unos 90,000 años, los Homo sapiens comenzaron a expandirse fuera de África, llegando a Oriente Medio y luego a Europa hace unos 45,000 años. En Europa, convivieron con los neandertales, quienes finalmente desaparecieron hace unos 28,000 años. Aunque se pensó que los neandertales eran una subespecie de Homo sapiens, los estudios genéticos sugieren que son especies diferentes, aunque hubo cruces entre ellos.
Homo sapiens: Nuestra especie
Nuestros parientes vivos más cercanos son los grandes simios: gorilas, chimpancés, bonobos y orangutanes. Los fósiles más antiguos de Homo sapiens tienen casi 200,000 años y se encontraron en Etiopía.
Los estudios genéticos indican que todos los humanos actuales descendemos de una misma "Eva mitocondrial", una mujer que vivió en el noreste de África hace entre 150,000 y 230,000 años. También hay un "Adán cromosómico" por línea paterna, que vivió en África hace entre 60,000 y 90,000 años.
Algunas teorías sugieren que la población de Homo sapiens se redujo drásticamente hace unos 74,000 años debido a la erupción de un supervolcán en Indonesia. Esto habría dejado a muy pocos individuos, lo que explicaría la gran similitud genética entre todos los humanos actuales.
En 2021, se descubrió una posible nueva especie de humano antiguo, el Homo longi o "Hombre dragón", a partir de un cráneo de hace unos 146,000 años encontrado en China. Los análisis sugieren que Homo sapiens está más relacionado con él que con los neandertales.
¿Cómo se expandió Homo sapiens por el mundo?
Los humanos comenzaron a salir de África hace unos 90,000 años.
- Australia y Nueva Guinea: Los humanos llegaron a Australia hace unos 50,000 años, usando balsas para cruzar el mar.
- Europa: Fue colonizada hace unos 40,000 años. La expansión por Europa coincidió con la desaparición de los neandertales.
- Oceanía: La colonización de las islas cercanas a Asia comenzó hace unos 50,000 años. Las islas más lejanas, como Hawái y Nueva Zelanda, se poblaron mucho después, hace unos 2,000 o 1,500 años, gracias al desarrollo de mejores barcos.
- América: Se cree que los humanos llegaron a América hace unos 20,000 o 15,000 años. Durante las épocas glaciales, el nivel del mar bajó, creando un puente de tierra (el Puente de Beringia) que unía Asia y América.
Cambios importantes en la evolución humana
Cambios en el cuerpo
¿Qué nos hace diferentes de otros primates?
Una diferencia clave es que los humanos perdimos la capacidad de producir vitamina C en nuestro cuerpo, algo que la mayoría de los animales sí pueden hacer. Esto pudo haber ocurrido cuando nuestros ancestros vivían en selvas con mucha fruta. Cuando las selvas se redujeron, tuvieron que buscar nuevas fuentes de alimento, lo que los obligó a moverse más.
Todos estos cambios han ocurrido en un tiempo relativamente corto (millones de años), lo que explica por qué nuestra especie puede tener problemas en la columna vertebral o en la circulación.
El desarrollo del cerebro
El cerebro del Homo sapiens es uno de los más grandes en proporción a nuestro cuerpo. Además, consume mucha energía: ¡el 20% de toda la energía del cuerpo!
El volumen del cerebro humano actual es de unos 1350 cm³. Los neandertales podían tener un cerebro más grande, pero su forma era diferente. El cráneo de Homo sapiens tiene una frente prominente y es más alto, lo que permite un mayor desarrollo de las partes frontales del cerebro, importantes para el pensamiento complejo.
Mutaciones en ciertos genes, como el gen FOXP2, fueron cruciales para el desarrollo de las áreas del cerebro relacionadas con el lenguaje. El cerebro humano también desarrolló la "lateralización", donde cada hemisferio tiene funciones especializadas (por ejemplo, el hemisferio izquierdo para el lenguaje).
Se cree que los Homo sapiens de hace 200,000 años ya tenían un potencial intelectual similar al nuestro. Sin embargo, las primeras expresiones artísticas y simbólicas (como dibujos en cuevas) aparecieron mucho después, hace unos 75,000 a 35,000 años. La escritura, una forma de "memoria" externa, surgió hace solo unos 5,500 años.
Nuestra cabeza es muy grande para contener este cerebro, lo que hace que los nacimientos sean difíciles. Por eso, los bebés humanos nacen muy inmaduros y necesitan muchos años de cuidado para desarrollarse completamente.
La bipedestación: Caminar sobre dos piernas
Los homininos, primates que caminan sobre dos patas, surgieron en África hace unos 6 o 7 millones de años. Una teoría es que esto fue una adaptación a la vida en la sabana, donde caminar erguido ayudaba a ver por encima de la vegetación y a evitar el sobrecalentamiento bajo el sol.
Para poder caminar erguidos, nuestro cuerpo sufrió grandes cambios:
- Cráneo: El orificio por donde la médula espinal se une al cerebro se movió hacia la base del cráneo.
- Columna vertebral: Adquirió curvas que actúan como un resorte y ayudan a soportar el peso del cuerpo.
- Pelvis: Se ensanchó para soportar mejor el peso de los órganos en posición vertical. Esto también hizo que el canal de nacimiento fuera más estrecho, dificultando los partos.
- Piernas: Se hicieron más fuertes y el fémur se inclinó hacia adentro, permitiendo una marcha más eficiente.
- Pies: Se alargaron, el dedo gordo dejó de ser oponible (como un pulgar) y se alineó con los demás, lo que nos ayuda a mantener el equilibrio y a impulsarnos al caminar.
Ventajas y desventajas de caminar erguido
Caminar sobre dos patas fue una gran ventaja en la sabana porque:
- Permitía ver a lo lejos para buscar comida o detectar depredadores.
- Liberaba las manos para transportar objetos (comida, herramientas, crías).
- Era más eficiente energéticamente para recorrer largas distancias.
- Reducía la exposición al sol y ayudaba a mantener el cuerpo fresco.
Sin embargo, también tuvo desventajas, como los partos más difíciles debido al cambio en la forma de la pelvis.
Manos libres: Una herramienta para la evolución
Al caminar erguidos, nuestras manos quedaron libres. Esto fue muy útil para recolectar alimentos como raíces, frutas, insectos y carroña. Se cree que nuestros ancestros eran carroñeros y actuaban en grupos para robar presas de otros animales.
Nuestras manos se especializaron. Aunque el pulgar oponible es una característica antigua de los primates, en los humanos se volvió mucho más preciso y delicado. Esto nos permitió fabricar herramientas, algo que Homo habilis ya hacía hace más de 2 millones de años.
La visión: Ver el mundo de otra manera
Los humanos heredamos de nuestros ancestros primates la visión en tres dimensiones y la capacidad de ver muchos colores. Nuestros ojos están al frente de la cabeza, lo que nos permite ver en profundidad. Esta visión aguda nos ayuda a observar el horizonte y es fundamental para nuestra forma de comunicarnos, por ejemplo, a través de gestos.
La especialización humana
A pesar de todos los cambios, Homo sapiens es una especie relativamente poco especializada. Muchos animales tienen adaptaciones muy específicas (como las pezuñas para correr), pero estas especializaciones pueden ser un riesgo si el ambiente cambia.
Nuestra falta de especializaciones anatómicas nos ha permitido adaptarnos a una gran variedad de ambientes. Además, los humanos tenemos características "juveniles" (neoténicas) en nuestra anatomía, como un rostro más plano y dientes pequeños. Esto nos hace vulnerables en la naturaleza, pero también nos impulsa a vivir en sociedades organizadas.
Aspectos culturales: El lenguaje y el pensamiento
El origen del lenguaje simbólico
El lenguaje humano, basado en símbolos y sonidos, es una de nuestras características más importantes. Para que pudiera surgir, se necesitaron cambios en la forma de nuestra boca y garganta. En los humanos, la laringe está más abajo que en otros mamíferos, lo que permite producir una mayor variedad de sonidos.
Los estudios en la Sierra de Atapuerca (España) sugieren que Homo antecessor, hace unos 800,000 años, ya tenía la capacidad de emitir un lenguaje oral. La fabricación de herramientas por Homo habilis hace 2 millones de años también sugiere que ya existía un lenguaje rudimentario para transmitir conocimientos.
La aparición del gen FOXP2 fue fundamental para el desarrollo del lenguaje y el pensamiento simbólico.
¿Cómo será el futuro de la evolución humana?
Hay diferentes ideas sobre cómo evolucionaremos en el futuro:
- Algunos creen que la evolución humana se ha ralentizado o detenido. Esto se debe a que los avances médicos permiten que sobrevivan personas que antes no lo harían, y la gran movilidad global mezcla los genes, reduciendo la "selección natural" tradicional.
- Otros piensan que la tecnología es ahora el motor de nuestra evolución. Por ejemplo, se dice que el entorno actual favorece la reproducción de personas inteligentes. Además, la ingeniería genética podría permitirnos elegir características para nuestros descendientes.
- También se ha propuesto que en el futuro, los humanos podrían vivir como "cyborgs" (parte humanos, parte máquinas) o incluso como seres digitales en cuerpos artificiales.
Galería de imágenes
-

Distribución geográfica y temporal del género Homo.
-

Excavación en el yacimiento de Gran Dolina, en Atapuerca (provincia de Burgos).
-

Comparación del esqueleto de simios y humanos por Thomas Henry Huxley.
-
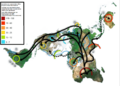
Mapa de la migración humana según estudios del ADN mitocondrial.




Véase también
 En inglés: Human evolution Facts for Kids
En inglés: Human evolution Facts for Kids
