
Stegoceras para niños
Datos para niños Stegoceras |
||
|---|---|---|
| Rango temporal: 77,5 Ma - 74 Ma Cretácico superior | ||
 Reconstrucción de S. validum, basada en partes de otros pequeños dinosaurios ornitisquios.
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Pachycephalosauria | |
| Familia: | Pachycephalosauridae | |
| Género: | Stegoceras Lambe, 1902 |
|
| Especie tipo | ||
| Stegoceras validum Lambe, 1902 |
||
| Otras Especies | ||
|
||
| Sinonimia | ||
|
||
Stegoceras es un género de dinosaurios paquicefalosáuridos. Vivieron al final del período Cretácico, hace unos 78 a 74 millones de años. Sus fósiles se han encontrado en lo que hoy es Norteamérica.
Este dinosaurio tenía dos especies conocidas. Los primeros restos se descubrieron en 1902 en Alberta, Canadá. La especie principal, Stegoceras validum, fue descrita a partir de estos hallazgos. En 2011, se nombró otra especie, Stegoceras novomexicanum, a partir de fósiles encontrados en Nuevo México.
Contenido
Stegoceras: El Dinosaurio con Cabeza Dura
Stegoceras era un dinosaurio pequeño que caminaba sobre dos patas. Medía entre 2 y 2,5 metros de largo y pesaba entre 10 y 40 kilogramos. Su característica más notable era su cráneo.
¿Cómo Era su Apariencia?

Stegoceras es uno de los paquicefalosáuridos mejor conocidos de Norteamérica. Su cráneo era más o menos triangular y tenía un hocico corto. La parte superior de su cabeza estaba cubierta por una cúpula gruesa y lisa. Esta cúpula se formaba por la unión de los huesos frontal y parietal.
El cráneo tenía una "plataforma" gruesa en la parte trasera, sobre la nuca. También tenía una cresta gruesa sobre los ojos. Gran parte del cráneo estaba decorado con pequeños bultos y nódulos. Algunos de estos formaban pequeños cuernos en la plataforma trasera.
Su Cráneo y Dientes
Los ojos de Stegoceras eran grandes y miraban un poco hacia adelante. Tenía aberturas nasales grandes orientadas hacia el frente. El hueso de la nariz era grueso y con una forma particular.
Sus dientes eran pequeños y tenían bordes aserrados, como una sierra. Eran diferentes entre sí y estaban bien sujetos en sus alvéolos. Tenía tres dientes en la parte delantera de la mandíbula superior, dieciséis en la parte trasera de la mandíbula superior y diecisiete en la mandíbula inferior. Los dientes de la parte delantera de la boca eran más puntiagudos. Los de la parte trasera eran triangulares.
Se cree que los cráneos de los Stegoceras jóvenes eran planos. Con la edad, estos cráneos se convertían en la cúpula redondeada que conocemos.
Su Esqueleto
La columna vertebral de Stegoceras era muy rígida. Esto se debía a que las articulaciones de sus vértebras impedían mucho el movimiento lateral. Además, tenía tendones que se habían vuelto hueso, lo que la hacía aún más fuerte.
Aunque no se conocen las vértebras de su cuello, se cree que lo mantenía en una postura curva, como una "S" o una "U". Esto es común en la mayoría de los dinosaurios. En su cola, tenía tendones osificados que formaban una especie de "canasta". Esto hacía que su cola fuera muy rígida.
La cintura pélvica de Stegoceras era muy ancha para un arcosaurio que caminaba sobre dos patas. Esta anchura podría haber ayudado a acomodar un intestino grande. Sus patas eran delgadas.
¿Cuándo y Dónde se Encontró?

Los primeros restos de Stegoceras fueron encontrados por el paleontólogo canadiense Lawrence Lambe en Alberta, Canadá, en 1898 y 1901. Eran principalmente cúpulas parciales de cráneos.
En 1902, Lambe describió y nombró a este nuevo dinosaurio como Stegoceras validus. El nombre Stegoceras viene del griego y significa "techo con cuerno", refiriéndose a su cabeza. El nombre validus significa "fuerte" en latín, quizás por la robustez de su cráneo.
Primeros Descubrimientos
Al principio, los científicos no estaban seguros de a qué grupo de dinosaurios pertenecía Stegoceras. Algunos pensaron que era pariente de los dinosaurios con cuernos o de los estegosáuridos (dinosaurios con placas).
En 1924, el paleontólogo estadounidense Charles W. Gilmore describió un cráneo completo de Stegoceras junto con otras partes del esqueleto. Este fue el hallazgo más completo de un dinosaurio con cabeza de cúpula hasta ese momento. Este descubrimiento ayudó a entender mejor a estos animales.
Cómo Cambió su Clasificación
Durante años, hubo mucha confusión sobre la clasificación de Stegoceras. Algunos científicos lo consideraron parte de otros géneros o lo agruparon con dinosaurios que no eran sus parientes cercanos.
Finalmente, en 1945, el paleontólogo estadounidense Charles M. Sternberg demostró que Stegoceras era diferente de otros dinosaurios. Propuso que los dinosaurios con cabeza de cúpula deberían tener su propia familia, a la que llamó Pachycephalosauridae. Stegoceras fue el primer miembro de esta familia en ser nombrado.
¿Cómo se Relaciona con Otros Dinosaurios?
En la década de 1970, se encontraron más fósiles de paquicefalosáuridos en Asia. Esto ayudó a los científicos a entender mejor a este grupo. En 1974, se concluyó que los paquicefalosáuridos eran lo suficientemente diferentes como para formar su propio suborden dentro de los Ornithischia.
En 1986, el paleontólogo estadounidense Paul Sereno propuso que los paquicefalosáuridos estaban relacionados con los ceratopsianos (dinosaurios con cuernos). Los unió en un grupo llamado Marginocephalia, basándose en características similares de sus cráneos.
El Árbol Familiar de los Paquicefalosáuridos
La clasificación de los paquicefalosáuridos se basa principalmente en las características de sus cráneos. Se dividían en grupos según si sus cúpulas eran planas o abovedadas. Sin embargo, más tarde se descubrió que las cabezas planas podían ser de animales jóvenes o una característica diferente, no siempre indicando un grupo distinto.
Los estudios más recientes sugieren que Stegoceras es uno de los paquicefalosáuridos más "básicos" o "primitivos". Esto significa que se encuentra en una rama temprana del árbol familiar de este grupo.
¿Cómo Vivía Stegoceras?
La vida de los paquicefalosáuridos, incluyendo a Stegoceras, aún tiene muchos misterios. Los fósiles nos dan pistas sobre cómo evolucionaron, qué comían y cómo usaban sus cabezas.
Cómo Comía
No se sabe con certeza qué comían los paquicefalosáuridos. Sus dientes eran pequeños y acanalados, lo que sugiere que no podían masticar plantas muy duras. Se cree que su dieta era variada, incluyendo hojas, semillas, frutas e insectos.
Los dientes de Stegoceras se parecían a los de algunos lagartos que comen plantas. La forma de su mandíbula sugiere que su boca se movía principalmente de arriba abajo, con poca capacidad de rotación. Esto indica que usaban la fuerza de su mordida más para cortar que para aplastar.
Cómo Crecía

Los científicos han estudiado cómo cambiaban los cráneos de Stegoceras a medida que crecían. Se ha propuesto que los cráneos planos que se encontraron eran de Stegoceras jóvenes. A medida que el dinosaurio envejecía, su cráneo se volvía más abovedado.
Los cráneos jóvenes tenían una superficie plana con pequeños bultos. En los subadultos, la cúpula empezaba a formarse. En los adultos, la cúpula era ancha y convexa, cubriendo la mayor parte de la parte superior del cráneo. Los bultos y nódulos se estiraban o desaparecían a medida que la cúpula crecía.
¿Para Qué Servía su Cúpula?

La función de la cúpula de Stegoceras ha sido muy debatida. Las teorías principales son que se usaba para:
- Combate entre individuos de la misma especie: Algunos científicos creen que la cúpula se usaba para golpear cabezas, como hacen algunas ovejas o cabras modernas. La cúpula era muy gruesa y fuerte, ideal para resistir impactos.
- Exhibición para atraer pareja o reconocimiento de especies: Otros sugieren que la cúpula era para mostrarse a otros de su especie, quizás para atraer pareja o para reconocerse entre ellos. Podría haber tenido colores brillantes.
Los estudios han simulado impactos en los cráneos de Stegoceras. Han encontrado que la cúpula podía soportar golpes considerables. El hueso esponjoso dentro de la cúpula podría haber protegido el cerebro. También se cree que la cúpula estaba cubierta de queratina, un material fuerte que ayuda a absorber la energía.
Se han encontrado lesiones en los cráneos de Stegoceras y otros paquicefalosáuridos. Estas lesiones, que parecen ser infecciones por golpes, se concentran en la parte superior de la cúpula. Esto apoya la idea de que se usaban para golpear cabezas en disputas por territorio o para competir por parejas.
Su Sentido del Olfato

Los estudios de los cerebros de Stegoceras sugieren que tenían un buen sentido del olfato. Esto se debe a que tenían grandes bulbos olfativos.
También se cree que tenían estructuras cartilaginosas en sus fosas nasales, llamadas cornetes nasales. Estas estructuras habrían ayudado a dirigir el aire hacia la región olfativa. Además, el sistema de vasos sanguíneos en los pasajes nasales sugiere que los cornetes también ayudaban a enfriar la sangre que iba al cerebro.
¿Dónde Vivía Stegoceras?
S. validum vivió en el Cretácico tardío en lo que hoy es Canadá. Sus fósiles se han encontrado en la Formación Dinosaur Park y la Formación Oldman en Alberta. S. novomexicanum se encontró en Nuevo México. Si esta especie pertenece a Stegoceras, el género habría vivido en una gran área geográfica.
Tradicionalmente, se pensaba que los paquicefalosáuridos vivían en zonas montañosas. Sin embargo, estudios más recientes sugieren que habitaban en llanuras aluviales y zonas costeras, cerca del agua.
La Formación Dinosaur Park, donde se encontraron muchos fósiles de Stegoceras, era un lugar con ríos y llanuras. El clima era más cálido que el actual, con estaciones húmedas y secas. En este lugar vivían muchos otros dinosaurios herbívoros, como Centrosaurus, Styracosaurus y Chasmosaurus. También había grandes depredadores como Gorgosaurus y Daspletosaurus.
Galería de imágenes
-

Molde del cráneo de S. validum UALVP 2, el primer cráneo completo de paquicefalosaurio.
-
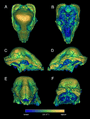
Imágenes de CT del cráneo UALVP 2 en vistas múltiples, que muestran las densidades superficiales relativas del hueso (azul: bajo, amarillo: alto).
-

Reconstrucción esquelética de Charles W. Gilmore de 1924 de UALVP 2, que muestra los tendones de la cola como gastralia.
-
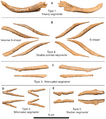
Tendones osificados de la cola de UALVP 2.
-

Dos esqueletos de S. validum reconstruidos basados en el espécimen UALVP 2, Royal Tyrrell Museum.
-

Cráneo parcial de un S. validum juvenil de cabeza plana, UCMZ(VP) 2008.001.
-

Ilustración de 1918 de la cúpula de S. validum CMN 138 en múltiples vistas.
-
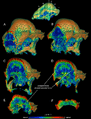
Imágenes de TC que muestran la densidad interna del hueso en UALVP 2 (azul: bajo, rojo: alto).
-
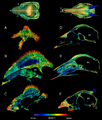
Cortes transversales de CT de los cráneos de UALVP 2 (izquierda) y un duiker de vientre blanco (derecha).
-

Restauración de un par con cúpulas de diferentes colores.
-
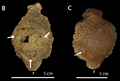
Dos cúpulas de S. validum con lesiones (flechas) mostradas desde arriba.
-

Imágenes de TC de TMP 84.5.1, un S. validum juvenil o subadulto, con secciones.
-
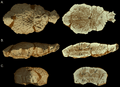
Imágenes de TC de AMNH 5450 juvenil de cabeza plana, "Ornatotholus", con secciones a la derecha.













Véase también
 En inglés: Stegoceras Facts for Kids
En inglés: Stegoceras Facts for Kids
- Anexo: Géneros válidos de dinosaurios
