
Dilophosaurus wetherilli para niños
Datos para niños Dilophosaurus wetherilli |
||
|---|---|---|
| Rango temporal: 183,7 Ma Jurásico Inferior | ||
|
Recreación
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Neotheropoda | |
| Superfamilia: | Coelophysoidea | |
| Familia: | Dilophosauridae | |
| Género: | Dilophosaurus Welles, 1970 |
|
| Especie: | D. wetherilli (Welles, 1954) |
|
| Sinonimia | ||
|
||

El Dilophosaurus wetherilli (que significa "lagarto de dos crestas de John Wetherill" en griego) es un dinosaurio terópodo que vivió hace unos 183 millones de años. Esto fue al principio del Jurásico, en lo que hoy es Norteamérica. Es la única especie conocida del género extinto Dilophosaurus.
Este dinosaurio medía unos 7 metros de largo y pesaba alrededor de 400 kilogramos. Era uno de los primeros grandes depredadores, aunque más pequeño que otros terópodos que vivieron después. Tenía un cuerpo delgado y ligero. Su cráneo era grande, pero delicado, con un hocico estrecho. Lo más llamativo eran dos crestas en forma de placa en su cabeza, parecidas a las de un casuario.
La mandíbula del Dilophosaurus era delgada en la parte delantera, pero más fuerte atrás. Sus dientes eran largos, curvos y delgados. Los del maxilar inferior eran más pequeños que los del superior. Tenía un cuello largo con vértebras huecas y ligeras. Sus brazos eran fuertes, con cuatro dedos en las manos. El primer dedo era corto y fuerte con una garra grande. Los dos siguientes eran más largos y delgados, y el cuarto era muy pequeño y no tenía función. Sus patas traseras eran robustas, con pies fuertes y grandes garras.
Los paleontólogos han estudiado mucho al Dilophosaurus. Algunos creen que es un ancestro de otros grupos de terópodos. Otros piensan que es un celofísido grande. Los primeros restos se encontraron en 1954, pero el nombre actual se le dio una década después. Es uno de los terópodos más antiguos del Jurásico, pero aún no se conoce del todo. Hoy se le considera parte de la familia Dilophosauridae, junto con el Dracovenator.
Se cree que el Dilophosaurus era un animal activo que caminaba sobre dos patas. Pudo haber cazado animales grandes y pequeños, incluyendo peces. La función de sus crestas no se sabe con certeza. Eran demasiado frágiles para pelear, pero quizás las usaba para reconocer a otros de su especie o para atraer pareja. Pudo haber crecido muy rápido, ganando entre 30 y 35 kilogramos al año al principio de su vida. Algunos fósiles muestran que tuvo lesiones y problemas de desarrollo. El Dilophosaurus se ha encontrado en la Formación Kayenta, donde vivía con otros dinosaurios como el Megapnosaurus y el Sarahsaurus.
El Dilophosaurus ha aparecido en la cultura popular, siendo su aparición más famosa en la película Parque Jurásico de 1993. En la película, se le muestra mucho más pequeño de lo que era en realidad. También se le dio la habilidad de desplegar un collar alrededor del cuello, como el lagarto Chlamydosaurus kingii, y de escupir veneno. Estas dos últimas características son inventos de la película y la novela en la que se basa, ya que no hay pruebas científicas de que el Dilophosaurus las tuviera. En 2017, fue nombrado el dinosaurio estatal de Connecticut.
Contenido
Características del Dilophosaurus
El Dilophosaurus fue uno de los primeros grandes dinosaurios carnívoros. Era un terópodo de tamaño mediano, aunque más pequeño que algunos terópodos posteriores. Su cuerpo era delgado y ligero, similar al tamaño de un oso pardo. El espécimen más grande conocido pesaba unos 400 kilogramos y medía unos 7 metros de largo. Su cráneo medía 590 milímetros. El primer espécimen encontrado, llamado holotipo, era más pequeño, con 6.03 metros de largo y 283 kilogramos de peso. Algunos científicos han encontrado huellas que sugieren que pudo haber tenido plumas suaves, como el plumón, alrededor de su vientre y patas.

La característica más notable del Dilophosaurus son las dos crestas redondeadas en su cráneo. Se cree que las usaba para exhibirse. Estas crestas aparecieron en el Dilophosaurus antes que en otros terópodos. Algunos estudios sugieren que pudo haber diferencias de tamaño entre machos y hembras. Los dientes del Dilophosaurus eran largos y delgados. Tenía una muesca detrás de la primera fila de dientes, lo que le daba un aspecto similar al de un cocodrilo. Esta muesca se debía a una conexión débil entre los huesos de la mandíbula superior. Al principio, se pensó que esto significaba que el Dilophosaurus era un carroñero, porque sus dientes delanteros parecían demasiado débiles para cazar presas grandes.
Estructura del cuerpo
El Dilophosaurus tenía 10 vértebras cervicales en el cuello, 14 en la espalda y 45 en la cola. Su cuello era largo y flexible, y sus vértebras eran muy ligeras y huecas. Esto hacía que su cuello fuera menos pesado. Los brazos eran fuertes y tenían huesos largos y delgados. Las manos tenían cuatro dedos. El primer dedo era corto pero fuerte, con una garra grande. Los dos siguientes eran más largos y delgados, con garras más pequeñas. El cuarto dedo era muy pequeño y no tenía una función clara.

Las patas traseras eran grandes y fuertes. El hueso del muslo era más largo que el de la parte inferior de la pierna. Tenía pies largos y robustos con tres dedos bien desarrollados que tenían grandes garras. El tercer dedo era el más fuerte. El primer dedo del pie era más pequeño y no tocaba el suelo.
Cráneo y mandíbula
El cráneo del Dilophosaurus era grande en proporción a su cuerpo, pero delicado. El hocico era estrecho. Las fosas nasales estaban más atrás que en la mayoría de los terópodos. Tenía un espacio entre los huesos delanteros de la mandíbula superior, lo que creaba una "muesca" en su fila de dientes.

Las dos crestas en la parte superior del cráneo eran altas, delgadas y arqueadas. Estaban formadas por los huesos lagrimales y nasales. Cada cresta tenía una pequeña proyección hacia atrás. Como solo se ha encontrado un espécimen con las crestas completas, no se sabe si variaban entre individuos.
La mandíbula era delgada en la parte delantera, pero fuerte donde se unía al cráneo. Tenía cuatro dientes en cada hueso premaxilar (delantero de la mandíbula superior), doce en cada maxilar (siguiente hueso de la mandíbula superior) y diecisiete en cada dentario (mandíbula inferior). Los dientes eran largos, delgados y curvos. Eran más grandes en la mandíbula superior. La mayoría de los dientes tenían pequeños bordes dentados en la parte delantera y trasera.

Descubrimiento e investigación
Los primeros restos de Dilophosaurus wetherilli fueron encontrados en 1942 por una expedición del Museo de Paleontología de la Universidad de California. Un hombre navajo llamado Jesse Williams les mostró unos huesos que había descubierto en 1940. Los fósiles se encontraron en la formación Kayenta, en el norte de Arizona. Se hallaron tres esqueletos de dinosaurios. El primero estaba casi completo.

El primer esqueleto fue limpiado y montado. En 1954, el paleontólogo Samuel P. Welles, quien participó en la excavación, describió el dinosaurio. Lo llamó Megalosaurus wetherilli, en honor a John Wetherill, un concejal navajo. En ese momento, muchos terópodos se agrupaban bajo el nombre Megalosaurus sin mucho orden.
En 1964, Welles regresó a la zona y encontró otro esqueleto, más grande. Durante su preparación, se hizo evidente que este dinosaurio tenía dos crestas en la parte superior del cráneo. Esto llevó a revisar el primer espécimen, donde también se encontraron restos de las crestas. Se concluyó que los dos primeros especímenes eran jóvenes, y el de 1964 era un adulto.
Welles se dio cuenta de que este dinosaurio no encajaba en el género Megalosaurus. En 1970, le dio el nuevo nombre de género Dilophosaurus, que significa "lagarto de dos crestas". Fue el primer terópodo conocido del Jurásico temprano y sigue siendo uno de los mejor conservados de esa época.
Nuevos hallazgos y huellas
En 2001, el paleontólogo Robert J. Gay identificó restos de al menos tres nuevos especímenes de Dilophosaurus. Uno de ellos era un individuo joven, el más joven conocido de este género.
Se han encontrado muchas huellas fósiles que se cree que fueron hechas por Dilophosaurus o dinosaurios similares. En 1971, Welles encontró huellas en la formación Kayenta y las llamó Dilophosauripus williamsi. Sin embargo, es difícil probar con certeza que una huella fue hecha por una especie específica de dinosaurio.
.jpg)
Algunas huellas encontradas en Massachusetts, llamadas Grallator minisculus, se interpretaron como la impresión de un dinosaurio sentado. Se observaron marcas que algunos científicos creen que podrían ser de plumas, lo que sugeriría que el Dilophosaurus pudo haber tenido plumas. Sin embargo, otros científicos piensan que estas marcas son solo efectos del barro.
Clasificación del Dilophosaurus
A lo largo de los años, el Dilophosaurus ha sido clasificado en varios grupos de terópodos. Al principio, se le consideró un celofisoideo grande. Sin embargo, estudios más recientes sugieren que el Dilophosaurus y otros "dilofosáuridos" están más relacionados con los terópodos tetanuranos que con los celofisoideos.

En 2005, se describió un nuevo género llamado Dracovenator de Sudáfrica, que se encontró estrechamente relacionado con Dilophosaurus. Esto sugirió que no pertenecían a los celofisoideos, sino a un grupo más "avanzado" llamado Neotheropoda.
En 2015, los paleontólogos Christophe Hendrickx y sus colegas definieron la familia Dilophosauridae para incluir a Dilophosaurus y Dracovenator. Esta familia parece ser un poco más "avanzada" que los celofisoideos. Comparten características con ambos grupos, lo que muestra su posición importante en el árbol genealógico de los dinosaurios.
Árbol familiar
Aquí puedes ver cómo se relaciona el Dilophosaurus con otros dinosaurios terópodos, según diferentes estudios:

| Theropoda |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| Neotheropoda |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
Cómo vivía el Dilophosaurus
Alimentación
Al principio, se pensó que el Dilophosaurus era un carroñero (que solo comía animales muertos) porque la conexión de sus huesos en la mandíbula parecía débil. Se creía que sus dientes eran demasiado frágiles para cazar presas grandes.
Sin embargo, otros paleontólogos como Robert T. Bakker y Gregory S. Paul no estaban de acuerdo. Ellos pensaron que el Dilophosaurus estaba bien adaptado para cazar animales grandes. Paul incluso dijo que los animales terrestres que solo comen carroña son un mito. Sugirió que el Dilophosaurus cazaba animales grandes como los prosaurópodos y que era muy capaz de matar animales pequeños.
Algunos científicos, como Milner y James I. Kirkland, han sugerido que el Dilophosaurus pudo haber comido pescado. Señalan que las puntas de sus mandíbulas se ensanchaban, formando una "roseta" de dientes entrelazados, similar a la de los espinosáuridos, que se sabe que comían pescado. Sus fosas nasales también estaban retraídas, lo que podría haber evitado que el agua salpicara sus narices al pescar. Además, tenían brazos largos con garras fuertes, útiles para atrapar peces.
Crecimiento y desarrollo
Welles pensó que los especímenes más pequeños de Dilophosaurus eran jóvenes y el más grande era un adulto. Otros científicos han sugerido que las diferencias de tamaño podrían deberse a que machos y hembras eran de tamaños distintos.

En 2005, Ronald S. Tykoski descubrió que la mayoría de los especímenes conocidos eran jóvenes, y solo el más grande era un adulto. Robert Gay no encontró pruebas de diferencias entre sexos en los esqueletos, pero no descartó que pudiera haberlas en las crestas.
Estudios de los huesos del Dilophosaurus muestran que crecía muy rápido. Pudo haber ganado entre 30 y 35 kilogramos al año en sus primeros años de vida. Los dientes de reemplazo crecían dentro de la mandíbula y salían cuando los viejos se caían.
Cómo se movía
Welles imaginó al Dilophosaurus como un animal muy activo, que caminaba sobre dos patas, como un avestruz grande. Sus brazos eran fuertes y flexibles, y no los usaba para caminar, sino para agarrar y cortar. Cuando se sentaba, se apoyaba en la parte inferior de su pelvis y en su cola.

Algunos científicos sugirieron que la pelvis del Dilophosaurus era débil, lo que podría indicar que vivía en el agua y era un buen nadador. Sin embargo, la fuerza de sus patas traseras sugiere que era ágil y rápido al caminar en tierra.
Estudios sobre el movimiento de sus brazos muestran que podía mover el húmero (hueso del brazo) hacia atrás, pero no mucho hacia adelante. Sus dedos no podían extenderse demasiado hacia atrás, pero sí resistir la dislocación al atrapar presas.
Las huellas de Dilophosauripus sugieren que el Dilophosaurus pudo haber viajado en grupos pequeños. Sin embargo, también es posible que las inundaciones arrastraran huesos de diferentes individuos y los depositaran juntos.
Se ha encontrado una huella de un Dilophosaurus en Utah que muestra al animal agachado. Esta huella es importante porque es la única que muestra claramente las impresiones de las manos de un terópodo. Esto nos da información valiosa sobre cómo usaban sus patas delanteras. La postura de cuclillas era muy similar a la de las aves modernas. También mostró que los primeros terópodos sostenían las palmas de sus manos hacia adentro, una hacia la otra.

Función de las crestas
Welles pensó que las crestas del Dilophosaurus podrían haberse usado para regular la temperatura de su cuerpo o para reconocer a otros de su especie. También podrían haber sido solo para adornarse. Los Czerkas señalaron que las crestas eran demasiado delicadas para usarse en peleas. Sugirieron que eran una forma de exhibición visual para atraer a una pareja.

En 2011, los paleontólogos Kevin Padian y John R. Horner propusieron que las "estructuras extrañas" en los dinosaurios, como las crestas, se usaban principalmente para el reconocimiento de especies. Sin embargo, otros científicos argumentaron que la selección de pareja era una explicación más probable, debido al alto costo de desarrollar estas estructuras.
Enfermedades y lesiones antiguas

Un espécimen de Dilophosaurus wetherilli muestra posibles daños en una vértebra y un posible absceso en un hueso del brazo. Otro ejemplar tenía un brazo izquierdo más pequeño de lo normal en comparación con un brazo derecho muy fuerte. Esto podría ser un signo de problemas de desarrollo.
Un espécimen del Museo de Paleontología de la Universidad de California (UCMP 37302) tiene ocho problemas diferentes en sus hombros y brazos. Incluyen tres tumores óseos, huesos rotos que sanaron en ambos brazos, dedos deformados y la incapacidad de usar una de sus patas delanteras. Se cree que muchas de estas heridas ocurrieron durante una pelea o un accidente. El dinosaurio debió haber sentido mucho dolor mientras se curaba.
En 2016, Senter y Sara L. Juengst examinaron estas lesiones. Descubrieron que este espécimen tenía el mayor número y variedad de enfermedades en los hombros y brazos de cualquier terópodo conocido. Todas las heridas se habían curado, lo que significa que el Dilophosaurus sobrevivió mucho tiempo después de estos eventos, quizás meses o años. El uso de sus patas delanteras para cazar debió verse afectado durante la curación. Esto sugiere que el dinosaurio pudo haber pasado mucho tiempo sin comer o haber subsistido con presas muy pequeñas que podía atrapar con la boca y los pies. El alto nivel de dolor que pudo haber experimentado también muestra que era un animal muy resistente.

Entorno antiguo
Los restos de Dilophosaurus se encontraron en la Formación Kayenta, en el noreste de Arizona. Esta formación se compone principalmente de rocas depositadas por ríos. Se cree que se formó durante las etapas Sinemuriano y Pliensbachiano del Jurásico Inferior, hace entre 196 y 183 millones de años.

La Formación Kayenta ha revelado muchos fósiles de organismos. La mayoría son de animales con columna vertebral. También se han encontrado restos de plantas, conchas de agua dulce y huellas de invertebrados.
Entre los animales con columna vertebral encontrados, hay tiburones, peces, salamandras, ranas, tortugas, lagartos, cocodrilos primitivos, pterosaurios, y varios tipos de dinosaurios. Además del Dilophosaurus, se han encontrado el Kayentavenator, el Megapnosaurus, el Sarahsaurus, el Scelidosaurus y el Scutellosaurus. También se han hallado fósiles de mamíferos primitivos.
Los científicos han encontrado varios lugares con muchos fósiles juntos en la Formación Kayenta. Esto incluye restos de tres tipos diferentes de celofisoideos, lo que muestra que había una gran variedad de estos dinosaurios.
Cómo se fosilizaron
Los esqueletos originales de Dilophosaurus se encontraron de lado, con la cabeza y el cuello curvados hacia atrás, una postura común en los fósiles de dinosaurios. Se cree que esta postura se debe a cómo el cuerpo se asentó en el sedimento después de la muerte.
Welles concluyó que los especímenes fueron enterrados donde murieron, sin ser arrastrados muy lejos. Sin embargo, el espécimen principal parecía haber sido movido por animales que comían carroña. Otros especímenes muestran señales de haber sido arrastrados por una corriente, quizás por una inundación.
El Dilophosaurus en la cultura popular

En la película Parque Jurásico, el Dilophosaurus es representado con una membrana retráctil en el cuello, similar a la de un clamidosaurio. También se le da la habilidad de escupir un veneno que ciega a sus víctimas. Estas características son inventos de la película y la novela, ya que no hay pruebas científicas de que el Dilophosaurus las tuviera. En la película, el Dilophosaurus también se muestra mucho más pequeño de lo que era en realidad.
Los productos relacionados con Parque Jurásico, como los videojuegos Jurassic Park: Operation Genesis y Jurassic World Evolution, también incluyen al Dilophosaurus. La película lo hizo famoso, aunque con algunas diferencias científicas.
En 2017, el Dilophosaurus fue nombrado el dinosaurio estatal de Connecticut. Esto se debe a que en 1966 se encontraron huellas que se cree que fueron hechas por un dinosaurio similar en Rocky Hill. Estas huellas, llamadas Eubrontes giganteus, ya eran el fósil estatal de Connecticut desde 1991. En 1981, se donó una escultura de Dilophosaurus al parque estatal de dinosaurios de Connecticut.
Galería de imágenes
-
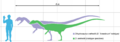
Tamaño de dos especímenes en comparación con un humano, con holotipo en verde y el espécimen más grande conocido en púrpura
-
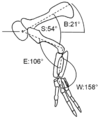
Diagrama que muestra la extremidad anterior de Dilophosaurus en la postura hipotética de reposo
-

Cráneo y cuello reconstruidos, Museo Real Tyrrell.
-

Cráneo reconstruido, mostrando las dos crestas que fueron la base del nombre Dilophosaurus, Museo Americano de Historia Natural
-

Réplica del holotipo con cráneo restaurado según el segundo espécimen, Museo Real Ontario, Toronto, Canadá
-

Esqueleto reconstruido basado en el holotipo, nótese la postura manual obsoleta
-

Huellas atribuidas en el Área de Recreación Nacional Glen Canyon , Arizona
-

Pista atribuida en Red Fleet Dinosaur Tracks Park, Utah
-

Esqueleto reconstruido de Sinosaurus sinensis, que originalmente fue descrito como una especie de Dilophosaurus, en MUSE, Trento
-

Restauración que muestra el tamaño de un adulto comparado con un humano.
-

Esqueleto de Coelophysis, con el que Dilophosaurus se agrupaba a menudo, Museo de Historia Natural de Cleveland.
-

Esqueleto reconstruido del terópodo crestado Cryolophosaurus, una vez considerado un pariente cercano de Dilophosaurus, Science World
-
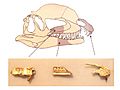
Cráneo de los huesos de Dracovenator, que puede ser el pariente más cercano de Dilophosaurus, en cuyo cráneo se trazan los huesos.
-
Réplica del cráneo y cuello, Museo Real Tyrrell.
-
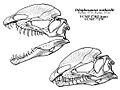
Diagramas de cráneo que muestran las mandíbulas abiertas y cerradas, así como la forma de cresta hipotéticamente reconstruida
-

Representación de un ambiente del Jurásico Inferior preservado en el Sitio de Descubrimiento de Dinosaurios St. George en Johnson Farm, con un Dilophosaurus wetherilli en una postura aviana de descanso
-

Huella de reposo del icnofósil Eubrontes, posiblemente asignable a Dilophosaurus, SGDS
-

Cabeza de una modelo apodada "Dyzio", Museo Geológico del Instituto Geológico del Estado, Varsovia
-
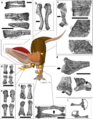
Patologías del espécimen UCMP 37302.
-

Restauración de la mano derecha del holotipo en flexión, con el tercer dedo deformado (debajo) incapaz de flexionar
-

Restauración de Dilophosaurus persiguiendo a Scutellosaurus.
-

Modelo de Dilophosaurus.




Véase también
 En inglés: Dilophosaurus Facts for Kids
En inglés: Dilophosaurus Facts for Kids
- Cryolophosaurus
- Parque Jurásico
- Jurásico
- Anexo: Géneros válidos de dinosaurios
