
Heterodontosaurus tucki para niños
Datos para niños Heterodontosaurus tucki |
||
|---|---|---|
| Rango temporal: 200 Ma - 190 Ma Jurásico Inferior | ||
 |
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Familia: | Heterodontosauridae | |
| Género: | Heterodontosaurus Charning & Crompton 1962 |
|
| Especie: | H. tucki Charning & Crompton 1962 |
|
| Sinonimia | ||
|
||
El Heterodontosaurus tucki es un dinosaurio que ya no existe. Su nombre significa "lagarto de dientes diferentes" en griego. Este dinosaurio vivió al principio del período Jurásico, hace unos 200 a 190 millones de años, en lo que hoy conocemos como África.
En la década de 1960, los científicos Alan Charning y Alfred Crompton encontraron los restos de este pequeño dinosaurio en una zona montañosa de Sudáfrica. Sus fósiles se hallaron en las formaciones geológicas llamadas Formación Elliot Superior y Formación Clarence.
Aunque era pequeño, el Heterodontosaurus era uno de los más grandes de su familia. Podía medir entre 1.20 y 1.75 metros de largo y pesar entre 2 y 10 kilogramos. Tenía un cuerpo corto con una cola larga. Sus patas delanteras tenían cinco dedos y eran fuertes, mientras que las patas traseras eran largas, delgadas y tenían cuatro dedos. Su cabeza era alargada y estrecha. La parte delantera de su boca tenía un pico duro. Lo más curioso eran sus dientes: tenía dientes pequeños como los de un conejo, seguidos de colmillos largos, y luego dientes con forma de cincel.
El Heterodontosaurus es el dinosaurio que da nombre a la familia Heterodontosauridae. Se cree que esta familia es una de las más antiguas dentro del grupo de los dinosaurios Ornithischia. A pesar de sus grandes colmillos, se piensa que el Heterodontosaurus comía plantas o, a veces, también pequeños animales. Aunque antes se pensaba que podía caminar en cuatro patas, ahora se cree que caminaba solo en dos.
Contenido
¿Cómo era el Heterodontosaurus?
El Heterodontosaurus era un dinosaurio de tamaño pequeño. El esqueleto más completo encontrado medía aproximadamente 1.18 metros de largo. Se ha calculado que pesaba entre 1.8 y 3.4 kilogramos. Un segundo cráneo incompleto sugiere que algunos Heterodontosaurus podrían haber sido más grandes, llegando a medir hasta 1.75 metros y pesar casi 10 kilogramos. No se sabe por qué había esta diferencia de tamaño, podría ser por variaciones normales o por diferencias físicas entre machos y hembras.
Este dinosaurio era uno de los más grandes de su familia, los Heterodontosauridae. Otros miembros de esta familia eran mucho más pequeños, como el Fruitadens de Norteamérica, que medía solo entre 65 y 75 centímetros.
Dientes y boca del Heterodontosaurus
El Heterodontosaurus tenía tres tipos de dientes diferentes. Tenía dientes pequeños y afilados, colmillos largos y curvos (que quizás solo tenían los machos), y dientes con forma de muela. Muchos fósiles de Heterodontosaurus no tenían colmillos. Al principio se pensó que eran individuos jóvenes, pero luego se vio que algunos adultos tampoco los tenían. Esto llevó a la idea de que las diferencias físicas entre machos y hembras podrían explicar la presencia o ausencia de colmillos.
En la parte delantera de su boca, tenía un pico. Detrás del pico, tenía dientes pequeños que probablemente usaba para cortar hojas y brotes de plantas.
Patas y cuerpo del Heterodontosaurus
Las patas traseras del Heterodontosaurus tenían tres dedos hacia adelante y uno hacia atrás. Este dedo trasero le ayudaba a mantener el equilibrio cuando estaba parado comiendo. Los huesos de su tobillo estaban unidos, de forma similar a los de las aves.
Sus muñecas eran pequeñas, pero muy flexibles. Esto le permitía agarrar y arrancar hojas. Cada mano tenía cinco dedos, y dos de ellos podían oponerse, como nuestro pulgar. Los primeros tres dedos terminaban en garras grandes y fuertes. Los dedos cuarto y quinto eran muy pequeños.
El Heterodontosaurus tenía una pelvis larga y estrecha. Su espalda y cola tenían varillas de hueso entrecruzadas que le daban soporte, en lugar de músculos pesados como los de dinosaurios más grandes.
Su cuello tenía 9 vértebras y formaba una curva en forma de "S". El tronco era corto, con 12 vértebras en la espalda y 6 unidas en la cadera. La cola era larga en comparación con el cuerpo, aunque no se conoce por completo, se cree que tenía entre 34 y 37 vértebras. La columna vertebral estaba rígida por tendones que se habían vuelto hueso, lo que ayudaba a soportar el cuerpo al caminar en dos patas. A diferencia de otros dinosaurios, su cola era flexible.

Las patas delanteras eran fuertes y largas, midiendo el 70% de la longitud de las patas traseras. La mano era grande y tenía cinco dedos adaptados para agarrar. Los primeros tres dedos terminaban en garras grandes y fuertes. Los dedos cuarto y quinto eran muy pequeños.
Las patas traseras eran largas y delgadas, terminando en cuatro dedos. El primer dedo no tocaba el suelo. Algo único en este tipo de dinosaurio es que varios huesos de la pierna y el pie estaban unidos, como en las aves modernas. Esto hacía que sus patas fueran muy eficientes para correr.
Algunos científicos creen que el Heterodontosaurus pudo haber tenido estructuras largas y filamentosas, como cerdas, desde el cuello hasta la cola, parecidas a las de un puercoespín.
Cráneo y dentadura del Heterodontosaurus
El cráneo del Heterodontosaurus era pequeño pero resistente. Medía entre 108 y 121 milímetros de largo. Era alargado, estrecho y triangular visto de lado. La parte más alta era una cresta en la parte superior de la cabeza. Los ojos eran grandes y redondos, con un hueso que sobresalía por encima. Debajo del ojo, tenía un hueso con forma de cuerno.
La parte delantera de la mandíbula no tenía dientes y estaba cubierta por un pico duro. Esto se sabe por las superficies rugosas en los huesos. El paladar (techo de la boca) era estrecho. Las aberturas de la nariz eran pequeñas.
Una característica inusual de su cráneo eran sus dientes de diferentes formas, algo que se ve más en los mamíferos. La mayoría de los dinosaurios tienen un solo tipo de diente. El Heterodontosaurus tenía tres tipos:
- La punta del pico no tenía dientes.
- Detrás del pico, en la mandíbula superior, tenía tres dientes pequeños y cónicos a cada lado, como incisivos.
- El tercer diente a cada lado era mucho más grande, formando colmillos prominentes.
- Los dos primeros dientes de la mandíbula inferior también eran colmillos, pero más grandes que los de arriba.
- Once dientes altos con forma de cincel se alineaban a cada lado de la parte trasera de la mandíbula superior, separados de los colmillos por un espacio grande. Estos dientes aumentaban de tamaño hacia el medio y luego disminuían. Tenían una capa gruesa de esmalte en el lado interno y estaban adaptados para masticar.
Los colmillos de la mandíbula inferior encajaban en un hueco en la mandíbula superior. Las filas de dientes superiores e inferiores encajaban bien, lo que creaba un "espacio para las mejillas" que también se ve en otros dinosaurios ornitisquios.
Descubrimiento e investigación del Heterodontosaurus

El primer fósil de Heterodontosaurus tucki, llamado SAM-PK-K337, fue encontrado durante una expedición en Sudáfrica y Lesoto entre 1961 y 1962. Hoy se encuentra en el Museo Iziko de Sudáfrica. Fue descubierto en una montaña a unos 1.890 metros de altitud.
Los científicos Alfred Walter Crompton y Alan J. Charig lo describieron y le dieron nombre en 1962. El nombre del género se refiere a sus dientes de diferentes formas, y el nombre de la especie, tucki, es en honor a George C. Tuck, quien apoyó la expedición. Este descubrimiento fue muy importante porque en ese momento se conocían pocos dinosaurios ornitisquios tempranos.
En 1966, se encontró un segundo fósil, SAM-PK-K1332, que incluía tanto el cráneo como el esqueleto, conservados en su posición natural. Este es el esqueleto de heterodontosáurido más completo encontrado hasta ahora. Aunque se prometió una descripción más detallada del cráneo, no se publicó hasta 2011, después de la muerte de Charig en 1997.
Otros fósiles de Heterodontosaurus incluyen un cráneo juvenil, partes de mandíbulas y dientes. En 2005, se encontró un nuevo fósil muy completo cerca de Grahamstown, pero las rocas que lo rodeaban eran muy duras. Por eso, en 2016, el fósil fue escaneado con tecnología avanzada para estudiar su anatomía.
En 1970, el científico Richard A. Thulborn sugirió que Heterodontosaurus era el mismo género que Lycorhinus, otro dinosaurio encontrado en Sudáfrica. Sin embargo, otros científicos no estuvieron de acuerdo, señalando que el fósil de Lycorhinus estaba muy fragmentado y era difícil de comparar. Finalmente, se decidió que eran géneros diferentes.
Clasificación del Heterodontosaurus
Cuando se describió en 1962, el Heterodontosaurus se clasificó como un miembro primitivo de los Ornithischia, uno de los dos grandes grupos de Dinosauria. Los científicos Alfred Romer y Oskar Kuhn nombraron la familia Heterodontosauridae en 1966, incluyendo al Heterodontosaurus y al Lycorhinus.
A principios del siglo XXI, las teorías más aceptadas eran que los heterodontosáuridos eran un grupo cercano a los Marginocephalia (que incluyen a los dinosaurios con cabeza gruesa y con cuernos) o a los Cerapoda (que incluyen a los ornitópodos). También se les considera uno de los grupos más antiguos de ornitisquios.
La familia Heterodontosauridae existió desde el Triásico superior hasta el Cretácico inferior, durante al menos 100 millones de años. Se han encontrado fósiles de heterodontosáuridos en África, Eurasia y América. La mayoría se han encontrado en el sur de África.
Se cree que los heterodontosáuridos se dividieron en dos grupos principales: uno con dientes de corona baja y otro con dientes de corona alta, como el Heterodontosaurus. Los de dientes de corona baja se encontraron en lo que fue Laurasia (el continente del norte), y los de dientes de corona alta en lo que fue Gondwana (el continente del sur).
¿Cómo se relaciona con otros dinosaurios?

Muchos géneros se han relacionado con la familia Heterodontosauridae, pero el Heterodontosaurus sigue siendo el más conocido. El siguiente diagrama muestra cómo se cree que se relacionan los miembros de esta familia: {{clade| style=font-size:100%;line-height:100% |label1=Heterodontosauridae |1=
|
|||||||||||||||||||||||||
| Heterodontosaurinae |
|
||||||||||||||||||||||||
¿Cómo vivía el Heterodontosaurus?
Crecimiento y desarrollo
No se sabe mucho sobre cómo crecía el Heterodontosaurus desde que era joven hasta que era adulto, porque se han encontrado pocos fósiles de juveniles. Sin embargo, el cráneo de un juvenil muestra que sus ojos se hacían más pequeños a medida que crecía, y su hocico se hacía más largo y le salían más dientes. La forma de sus dientes no cambiaba con la edad, lo que sugiere que los jóvenes y los adultos comían lo mismo.
¿Tenían diferencias físicas entre machos y hembras?
En 1974, se sugirió que los grandes colmillos del Heterodontosaurus eran una característica que solo tenían los machos adultos. Se pensó que el Abrictosaurus, un pariente cercano que no tenía colmillos, podría haber sido una hembra.
Sin embargo, en 2006, otros científicos encontraron que el cráneo de un Heterodontosaurus juvenil ya tenía colmillos. Esto hizo dudar de la idea de que los colmillos fueran solo de los machos adultos. Además, casi todos los Heterodontosaurus conocidos tienen colmillos, lo que no encaja con la idea de que la mitad fueran machos con colmillos y la otra mitad hembras sin ellos.
¿Cómo cambiaban sus dientes?
Hubo mucho debate sobre si el Heterodontosaurus cambiaba sus dientes continuamente, como otros dinosaurios y reptiles. Algunos estudios iniciales sugirieron que no, y que todos sus dientes se gastaban por igual, lo que indicaba que se formaban al mismo tiempo. Esto llevó a la idea de que el Heterodontosaurus podría haber reemplazado todos sus dientes a la vez, quizás durante un período de inactividad (como una especie de hibernación) en épocas secas.
Sin embargo, estudios posteriores mostraron que los patrones de desgaste de los dientes indicaban movimientos de mandíbula verticales y laterales, y que sí había variaciones en el desgaste, lo que sugería un reemplazo continuo de los dientes. Se descubrió que los dientes de reemplazo estaban presentes incluso en los juveniles. Esto significa que el Heterodontosaurus reemplazaba sus dientes de forma esporádica, no continuamente como otros dinosaurios.
¿Cómo se movía y qué comía?

Aunque la mayoría de los científicos creen que el Heterodontosaurus era un corredor que caminaba en dos patas, algunos estudios anteriores sugirieron que podía moverse en cuatro patas. Sus brazos eran fuertes y robustos, lo que podría haberle servido para desenterrar raíces o romper nidos de insectos.
Se cree que los dinosaurios eran animales de sangre caliente, con un metabolismo alto, similar al de los mamíferos y aves actuales. Un estudio de 2009 sugirió que el Heterodontosaurus habría necesitado un metabolismo alto para correr, lo que indica que era de sangre caliente.
El Heterodontosaurus es considerado generalmente un dinosaurio que comía plantas. En 1974, se propuso que sus colmillos no eran para comer, sino para pelear con otros de su especie, para mostrarse o para defenderse. Funciones similares se ven en los colmillos de algunos ciervos pequeños actuales.

Estudios más recientes han planteado la posibilidad de que fuera un omnívoro y usara sus colmillos para cazar ocasionalmente. Algunos científicos notaron que la forma de sus dientes y colmillos recordaba a los animales que comen carne. Sin embargo, otros argumentaron que si los colmillos fueran para pelear, solo los tendrían los machos, y como se encuentran en casi todos los individuos, es más probable que tuvieran una función relacionada con la alimentación o la defensa.
En 2011, se señaló que sus brazos y manos eran largos y tenían garras grandes, y sus patas traseras le permitían correr rápido. Estas características podrían haberle permitido atrapar presas pequeñas. Como omnívoro, el Heterodontosaurus habría tenido una ventaja en la estación seca cuando la vegetación era escasa.
En 2012, el científico Sereno señaló varias características de su cráneo y dientes que sugieren una dieta principalmente de plantas. Tenía un pico duro y mejillas especializadas para cortar vegetación. Los músculos de su mandíbula eran grandes, y la articulación de la mandíbula estaba por debajo del nivel de los dientes, lo que permitía una mordida uniforme para masticar plantas. Se cree que los heterodontosáuridos eran como los pecaries de hoy, que tienen colmillos similares y comen una variedad de plantas como raíces, frutas y semillas.
¿Dónde vivía el Heterodontosaurus?

Los fósiles de Heterodontosaurus se han encontrado en las formaciones geológicas Formación Elliot Superior y Formación Clarens en el Supergrupo Karoo de Sudáfrica. Estas formaciones datan del Jurásico inferior, hace unos 200-190 millones de años.
La Formación Elliot Superior tiene rocas de color rojo/púrpura y arena roja/blanca. La Formación Clarens, un poco más joven, tiene arena de color crema/blanco. Ambas formaciones son famosas por sus muchos fósiles de animales, incluyendo anfibios, tortugas, cocodrilos primitivos y otros reptiles.
Otros dinosaurios encontrados en estas formaciones incluyen el Lesothosaurus, el Massospondylus y el Megapnosaurus. La Formación Elliot Inferior tiene la mayor diversidad de heterodontosáuridos conocida, incluyendo Heterodontosaurus, Lycorhinus, Abrictosaurus y Pegomastax.
La gran variedad de heterodontosáuridos en la misma zona sugiere que diferentes especies podrían haber comido distintos tipos de alimentos para evitar competir entre sí. Por ejemplo, el Heterodontosaurus, con sus dientes especializados, podría haber comido plantas más duras, mientras que el Abrictosaurus, menos especializado, podría haber comido vegetación más blanda.
Galería de imágenes
-

Heterodontosaurus comparado en tamaño con un humano.
-

Cráneo del molde de SAM-PK-K1332 y reconstrucción del diagrama de cráneo.
-
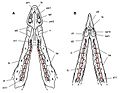
Diagramas que muestran la dentición de la mandíbula superior e inferior.
-
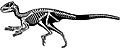
Diagrama esquelético de SAM-PK-K1332.
-
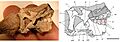
AMNH 24000, una calavera parcial.
-
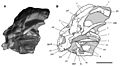
SAM-PK-K10487, un cráneo juvenil.
-
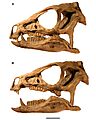
Cráneo restaurado.
-
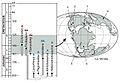
Distribución geográfica de heterodontosáuridos a lo largo del tiempo.
-

Modelo, que muestra una estructura de visualización hipotética en el hocico y cráneo.
-
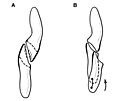
Diagrama que muestra el desgaste de la corona y la oclusión entre un diente superior e inferior de la mejilla en una vista lateral.









Véase también
 En inglés: Heterodontosaurus Facts for Kids
En inglés: Heterodontosaurus Facts for Kids
