
Dimetrodon para niños
Datos para niños Dimetrodon |
||
|---|---|---|
| Rango temporal: 295 Ma - 272 Ma Pérmico Inferior | ||
 Esqueleto de la especie Dimetrodon incisivum.
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Synapsida | |
| Orden: | Pelycosauria | |
| Suborden: | Eupelycosauria | |
| (sin rango): | Sphenacodontia | |
| Familia: | Sphenacodontidae | |
| Género: | Dimetrodon Cope, 1878 |
|
| Especie tipo | ||
| Dimetrodon limbatus (Cope, 1877 [originalmente Clepsydrops limbatus]) |
||
| Otras Especies | ||
| Sinonimia | ||
|
Bathyglyptus Case, 1911 |
||
El Dimetrodon (que significa "diente de dos medidas") es un género de animales extintos que vivieron hace mucho tiempo. Pertenecía a un grupo llamado sinápsidos y vivió en lo que hoy es América del Norte y Europa. Esto fue durante el Pérmico Inferior, hace entre 295 y 272 millones de años.
Lo más llamativo del Dimetrodon era su enorme vela en la espalda, parecida a una aleta. Esta vela estaba hecha de espinas largas que salían de su columna vertebral. Se cree que esta vela pudo haber ayudado al animal a controlar la temperatura de su cuerpo, calentándose o enfriándose. También podría haber servido para atraer a otros Dimetrodon durante el apareamiento.
Este animal era un depredador muy importante en su época. Se alimentaba de peces y otros animales de cuatro patas, como reptiles y anfibios. Caminaba sobre sus cuatro patas y tenía una cabeza grande con dientes de diferentes tamaños. La mayoría de los fósiles de Dimetrodon se han encontrado en el suroeste de Estados Unidos. Recientemente, se descubrió una nueva especie en Alemania. Esto no es tan raro, ya que en el Pérmico, América del Norte y Europa estaban unidas en un supercontinente llamado Euramerica.
Aunque el Dimetrodon se parece a un reptil y a menudo se confunde con un dinosaurio, en realidad vivió unos 40 millones de años antes de que aparecieran los primeros dinosaurios. Es más, el Dimetrodon está más relacionado con los mamíferos que con los reptiles modernos. Sin embargo, no es un antepasado directo de los mamíferos que conocemos hoy.
Contenido
Historia de los Descubrimientos

Primeras Investigaciones en el Siglo XIX
Los primeros estudios sobre los fósiles de Dimetrodon los hizo un paleontólogo estadounidense llamado Edward Drinker Cope en la década de 1870. Cope consiguió estos fósiles, junto con muchos otros restos de animales del Pérmico, de personas que los encontraron en una zona de Texas conocida como los Lechos Rojos.
Otros científicos también encontraron fósiles de Dimetrodon. Por ejemplo, Charles Hazelius Sternberg envió algunos a un paleontólogo alemán, Ferdinand Broili. El rival de Cope, Othniel Charles Marsh, también obtuvo algunos fósiles. El nombre Dimetrodon se usó por primera vez en 1878, cuando Cope describió varias especies.
Sin embargo, la primera vez que se describió un fósil de Dimetrodon fue un año antes, en 1877. Cope lo llamó Clepsydrops limbatus. Más tarde, en 1940, se confirmó que este fósil era en realidad un Dimetrodon.
Cope también fue el primero en describir un sinápsido con una vela en la espalda en 1878. Al principio, pensó que la vela era como una aleta, similar a la cresta de los basiliscos actuales.
Estudios a Principios del Siglo XX

A principios del siglo XX, el paleontólogo estadounidense E. C. Case realizó muchos estudios sobre el Dimetrodon. Él describió varias especies nuevas. Case recibió fondos para estudiar muchos fósiles que estaban en museos, algunos de los cuales habían sido encontrados por Cope.
En la década de 1920, otro paleontólogo, Alfred Romer, revisó muchos fósiles de Dimetrodon y describió más especies. En 1940, Romer y Llewellyn Ivor Price publicaron un estudio muy importante donde revisaron todas las especies de Dimetrodon conocidas. La mayoría de las especies que ellos consideraron válidas siguen siéndolo hoy.
Nuevos Hallazgos de Fósiles
Después de los estudios de Romer y Price, se encontraron nuevos fósiles de Dimetrodon en lugares fuera de Texas y Oklahoma. Se descubrieron en Utah en 1966, en Arizona en 1969 y en Ohio en 1975. Una nueva especie, D. occidentalis, se describió en 1977 en Nuevo México.
Antes de estos hallazgos, se pensaba que una parte del océano había separado a los Dimetrodon en una pequeña región de América del Norte. Sin embargo, los nuevos descubrimientos demostraron que este océano no era una barrera tan grande.
En 2002, se encontró una nueva especie de Dimetrodon, llamada D. teutonis, en Alemania. Este fue el primer hallazgo de este animal fuera de América del Norte.
¿Cómo se Relaciona con Otros Animales?

El Dimetrodon es un sinápsido primitivo. Los sinápsidos son un grupo diverso que incluye a los mamíferos de hoy y a todos sus parientes extintos. Aunque a menudo se confunde con un dinosaurio, el Dimetrodon apareció al menos 40 millones de años antes que los primeros dinosaurios.
De hecho, el Dimetrodon está más emparentado con los mamíferos actuales que con los dinosaurios o cualquier reptil moderno. Los científicos solían clasificar al Dimetrodon como un reptil. Sin embargo, ahora sabemos que los mamíferos evolucionaron de un grupo de animales relacionados con el Dimetrodon.
Los científicos dividen a los descendientes del ancestro común de mamíferos y reptiles en dos grandes grupos:
- Los sinápsidos: que incluyen al Dimetrodon y a los mamíferos.
- Los saurópsidos: que incluyen a los reptiles y aves de hoy, y a todos los reptiles extintos (como los dinosaurios).
Dentro de los sinápsidos, el Dimetrodon pertenece a un grupo llamado Sphenacodontia. Este grupo incluye al Dimetrodon y otros sinápsidos con velas dorsales, como el Secodontosaurus. También incluye a los terápsidos, que son un grupo más avanzado de sinápsidos que dieron origen a los mamíferos.
¿Cómo era el Dimetrodon?


El Dimetrodon era un animal de cuatro patas con una gran vela en la espalda. Su tamaño y forma eran similares a los de un caimán moderno, pero vivía solo en tierra. Fue el depredador principal de su ambiente durante más de 25 millones de años.
Había muchas especies de Dimetrodon, y con el tiempo se hicieron más grandes. La mayoría medía entre 1.7 y 3.2 metros de largo, y pesaba entre 28 y 250 kilogramos. La especie más grande, D. grandis, medía 3.2 metros. La más pequeña, D. teutonis, solo medía 60 cm.
Algunas características que lo relacionan más con los mamíferos que con los reptiles son: una mandíbula inferior fuerte, dientes de diferentes tamaños para masticar, músculos de la mandíbula especiales, un cerebro más grande y tres pequeños huesos en el oído medio.
Su Cabeza

El cráneo del Dimetrodon era alto, estrecho y triangular visto desde arriba. Tenía agujeros para los ojos en la parte superior y trasera. También tenía otros agujeros en la parte posterior del cráneo. La parte de atrás de su cabeza estaba un poco inclinada hacia arriba.
Visto de lado, la parte superior de su cráneo se curvaba hacia abajo hasta la punta del hocico. La punta de su mandíbula superior sobresalía un poco, creando un espacio entre sus dientes.
Los músculos de su mandíbula eran parecidos a los de los reptiles modernos. Esto, junto con sus dientes, sugiere que el Dimetrodon podía cazar y comer animales grandes.
Sus Dientes
Los dientes del Dimetrodon eran de diferentes tamaños, lo que le dio su nombre: "diente de dos medidas". Tenía uno o dos pares de dientes grandes, como colmillos, en la mandíbula superior. También tenía incisivos grandes en la punta de ambas mandíbulas. Los dientes más pequeños estaban detrás de los colmillos y se hacían más pequeños hacia la parte trasera de la boca.
Muchos de sus dientes eran más anchos en el medio y se estrechaban hacia la mandíbula, dándoles una forma ovalada. Esta forma es única del Dimetrodon y sus parientes cercanos. Como otros sinápsidos primitivos, la mayoría de los dientes del Dimetrodon tenían bordes aserrados, como una sierra.
Su Nariz
Dentro de la nariz del Dimetrodon, los fósiles muestran unas crestas que pudieron haber sostenido el tejido para detectar olores. Estas crestas eran más pequeñas que las de los sinápsidos más recientes. En esos animales más recientes, las crestas grandes sugieren que eran de sangre caliente, ya que ayudaban a calentar y humedecer el aire que respiraban. Por eso, la nariz del Dimetrodon es como un paso intermedio entre los primeros animales terrestres y los mamíferos.
Mandíbula y Oído
Otra característica interesante del Dimetrodon es una cresta en la parte superior de su mandíbula inferior. Esta cresta estaba en el hueso que conectaba la mandíbula con el cráneo. En los antepasados de los mamíferos, este hueso se separó de la mandíbula y se convirtió en el martillo, uno de los pequeños huesos del oído medio. La cresta del Dimetrodon se parece a una estructura que soporta el tímpano en los mamíferos.
Su Cola

La cola del Dimetrodon era muy larga y estaba formada por unas 50 vértebras. Al principio, los científicos pensaban que tenía una cola corta porque los primeros fósiles estaban incompletos. Pero en 1927, se encontró un fósil con una cola casi completa, lo que cambió esa idea.
La Vela Dorsal

La característica más famosa del Dimetrodon es su gran vela en la espalda. Esta vela estaba hecha de espinas vertebrales muy largas. Cada espina tenía una forma especial que la hacía fuerte y evitaba que se doblara o rompiera.
La parte de la espina más cercana al cuerpo era rugosa, lo que sugiere que allí se unían los músculos de la espalda. La parte más lejana era más lisa y tenía pequeños surcos, probablemente para los vasos sanguíneos que alimentaban la vela.
En algunos fósiles, se ven espinas que se curvan bruscamente. Esto podría significar que la vela no llegaba hasta la punta de las espinas, y que el tejido entre ellas no era tan extenso como se pensaba.
¿Para qué servía la Vela?

Hay muchas ideas sobre la función de la vela dorsal. Al principio, se sugirió que podría haber servido para camuflarse o incluso para impulsarse en el agua. Otra idea era que ayudaba a estabilizar el cuerpo al caminar.
En 1940, Alfred Romer y Llewellyn Ivor Price propusieron que la vela ayudaba al Dimetrodon a regular su temperatura corporal, calentándose con la luz del sol. Se hicieron estudios que calcularon cuánto tiempo le tomaba calentarse. Un estudio de 1986 sugirió que la aleta le permitía calentarse rápido por la mañana, pero no era tan buena para enfriarse o mantenerse caliente por la noche. Otro estudio de 1999 concluyó que la vela era útil para capturar y liberar calor durante todo el año.
Se cree que el Dimetrodon, como otros animales terrestres del Pérmico, era de sangre fría (ectotérmico), lo que significa que dependía del sol para mantener su temperatura. Como era grande, tardaba más en calentarse que sus presas más pequeñas. La vela podría haberle ayudado a calentarse más rápido por la mañana para poder cazar. También podría haberle ayudado a liberar el exceso de calor.

Estudios más recientes sugieren que la vela no era muy eficiente para regular la temperatura. La presencia de velas en especies pequeñas de Dimetrodon no apoya la idea de la termorregulación, ya que los cuerpos pequeños pueden calentarse y enfriarse fácilmente sin una vela grande. Además, parientes cercanos del Dimetrodon, como el Sphenacodon, tenían crestas dorsales pequeñas que no servían para regular la temperatura. Esto sugiere que la vela grande del Dimetrodon evolucionó por otras razones.
¿Para Mostrar?

Los Dimetrodon más grandes tenían las velas más grandes en proporción a su tamaño. Un estudio de 2010 encontró que la vela crecía más rápido de lo necesario para la termorregulación. Los investigadores concluyeron que la razón principal de la evolución de esta vela fue la selección sexual. Esto significa que la vela pudo haber sido usada para atraer a una pareja, como una forma de "mostrar" su fuerza o salud.
Diferencias entre Machos y Hembras


Es posible que el Dimetrodon tuviera dimorfismo sexual, lo que significa que había diferencias de tamaño entre machos y hembras. Algunos fósiles se consideran machos porque tienen huesos más gruesos, velas y cráneos más grandes, y mandíbulas más anchas. Otros fósiles, con características más pequeñas, podrían ser hembras.
¿Dónde Vivía y Qué Comía?

Los fósiles de Dimetrodon se han encontrado en América del Norte (Estados Unidos) y Europa (Alemania). Estas regiones formaban parte del supercontinente Euramérica en el Pérmico. La mayoría de los fósiles provienen de los "Lechos Rojos" de Texas y Oklahoma.
Estos lugares eran tierras bajas y planas, posiblemente grandes pantanos, durante el Pérmico. Los Lechos Rojos de Texas son famosos por la gran cantidad de fósiles de animales de cuatro patas. Además del Dimetrodon, allí vivían anfibios como el Eryops y reptiles como el Captorhinus.
Su Cadena Alimenticia
El Dimetrodon era un superdepredador en su ecosistema. Probablemente cazaba una variedad de animales, incluyendo peces, anfibios como el Trimerorhachis y el Diplocaulus, y otros animales terrestres como el Seymouria. También había insectos y pequeños reptiles que podrían haber sido parte de su dieta.
En los Lechos Rojos también vivían grandes herbívoros terrestres, como el Edaphosaurus. Estos animales se alimentaban de plantas terrestres y no dependían del agua para sobrevivir. Algunos científicos comparan el ecosistema del Dimetrodon con los Everglades de hoy.
La especie Dimetrodon teutonis, encontrada en Alemania, era más pequeña (alrededor de 1.7 metros). Es probable que se alimentara de invertebrados pequeños e insectos, ya que era demasiado pequeña para cazar a los grandes herbívoros de la zona. A diferencia de los pantanos de Texas, el lugar donde se encontró D. teutonis era un ambiente montañoso sin especies acuáticas.
Crías de Dimetrodon
Aunque algunas especies de Dimetrodon eran muy grandes, también se han encontrado muchos fósiles de crías. Un paleontólogo sugirió en 1982 que los adultos vivían en llanuras inundadas, mientras que los jóvenes preferían pantanos aislados y las orillas de los lagos.
Se pensaba que las crías de Dimetrodon se comportaban como las crías de los reptiles grandes de hoy, evitando a los adultos de su propia especie. Esto se debe a que los adultos podrían competir por la comida o incluso cazar a los jóvenes. Sin embargo, un estudio posterior puso en duda esta idea, sugiriendo que los fósiles de diferentes tamaños encontrados en esos hábitats eran de especies distintas, no de adultos y jóvenes de la misma especie.
Especies de Dimetrodon
Se han descrito muchas especies de Dimetrodon a lo largo de los años. Aquí te mostramos algunas de las más conocidas:
- Dimetrodon angelensis
- Dimetrodon dollovianus
- Dimetrodon giganhomogenes
- Dimetrodon grandis
- Dimetrodon limbatus
- Dimetrodon loomisi
- Dimetrodon macrospondylus
- Dimetrodon milleri
- Dimetrodon natalis
- Dimetrodon occidentalis
- Dimetrodon teutonis
Galería de imágenes
-

Recreación de Charles R. Knight, un Dimetrodon (frente) y un Edaphosaurus (atrás) basado en los especímenes de Cope.
-
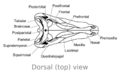
Huesos del cráneo de Dimetrodon (vista dorsal)
-
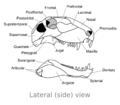
Huesos del cráneo de Dimetrodon (vista lateral)
-

Huesos del cráneo de Dimetrodon (vista ventral)
-

Huesos del cráneo de Dimetrodon (vista occipital)
-

Esqueleto AMNH 4636, posible macho.
-

Esqueleto del Museo de Historia Natural de la Universidad de Míchigan, posible hembra.





Véase también
 En inglés: Dimetrodon Facts for Kids
En inglés: Dimetrodon Facts for Kids
