
Chromista para niños
Datos para niños Chromista |
||
|---|---|---|
 En el sentido de las agujas del reloj desde la esquina superior izquierda: haptofita, diatomeas, oomiceto, criptofitas y alga parda.
|
||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| (sin rango) | Corticata | |
| Reino: | Chromista (P?) Caval.-Sm. 1981 |
|
| Subdivisiones | ||
|
||
Los cromistas son un grupo de seres vivos eucariotas, lo que significa que sus células tienen un núcleo definido. Algunos científicos los consideran un reino biológico aparte. Este grupo incluye a la mayoría de las algas que tienen clorofilas a y c en sus cloroplastos, que son como las "fábricas de alimento" de las células.
Los cloroplastos de los cromistas suelen estar rodeados por cuatro membranas. Se cree que los obtuvieron de un alga roja en el pasado. Algunos cromistas también tienen clorofilas a y b, que podrían venir de un alga verde. Además, hay cromistas que no tienen cloroplastos, pero están muy relacionados con los que sí los tienen.
Entre los cromistas más conocidos están las diatomeas (algas unicelulares), los dinoflagelados, las algas doradas, las haptofitas y las criptofitas. También se incluyen las algas pardas, que son pluricelulares. Algunos cromistas que no hacen fotosíntesis son los ciliados, foraminíferos, radiolarios, heliozoos y los oomicetos.
Contenido
¿Qué son los Cromistas?
El nombre Chromista fue propuesto por el científico Thomas Cavalier-Smith en 1981. Él quería agrupar a los Heterokontophyta (heterocontos), Haptophyta (haptofitas) y Cryptophyta (criptofitas). Otros nombres anteriores, como Chromophyceae o Chromophyta, se referían a grupos similares.
Más tarde, en 2010, Cavalier-Smith amplió la definición de Chromista para incluir a los Rhizaria y Centrohelida. En la clasificación de 2015, el reino Chromista abarca varios grupos importantes:
- Heterokonta
- Alveolata
- Rhizaria (estos tres forman el subreino Harosa)
- Haptophyta
- Cryptophyta
- Otros organismos heterótrofos relacionados (estos últimos en el subreino Hacrobia)
Los científicos han tenido dificultades para entender cómo se relacionan todos estos grupos entre sí. Sin embargo, estudios recientes sugieren que Heterokontophyta, Alveolata y Rhizaria están relacionados y forman un grupo llamado supergrupo SAR.
Características de los Cromistas
Históricamente, muchos cromistas fueron confundidos con plantas debido a sus paredes celulares con celulosa y su capacidad de hacer fotosíntesis. Otros, como los apicomplejos y ciliados, se consideraban protozoos. Los oomicetos incluso se pensaban que eran hongos.
Aunque algunos cromistas no pueden hacer fotosíntesis, la mayoría sí son autótrofos (producen su propio alimento). Los cromistas fotosintéticos suelen tener clorofilas a y c, además de otros pigmentos. Una excepción son las algas clorarácneas, que tienen clorofilas a y b.
Los cromistas no tienen muchas características físicas en común que se vean en todos ellos. Cada subgrupo tiene sus propias características únicas, como los alvéolos en Alveolata o los dos flagelos diferentes en Heterokontophyta.
Las características que sí comparten la mayoría de los cromistas son:
Fotosíntesis: ¿Cómo Producen su Alimento?

Una característica importante de muchos cromistas es su capacidad para hacer fotosíntesis. Se cree que sus plastos (los orgánulos que hacen la fotosíntesis) se originaron cuando una célula cromista "capturó" a otra alga (generalmente un alga roja) y la incorporó a su interior. Este proceso se llama endosimbiosis.
Como resultado de estas "capturas", los cloroplastos de los cromistas suelen tener cuatro membranas. Esto es diferente de las plantas, que tienen cloroplastos con solo dos membranas.
Cubierta Celular: Su Protección

La mayoría de los cromistas tienen algún tipo de cubierta celular. Esta cubierta les da una forma definida, cierta rigidez y los protege del exterior. Muchos tienen celulosa en sus paredes celulares, similar a las plantas.
Algunos cromistas tienen cubiertas muy complejas, como exoesqueletos, conchas o caparazones. Estas estructuras han dejado muchos microfósiles que los científicos estudian.
Importancia de los Cromistas en la Naturaleza


Los cromistas juegan un papel muy importante en nuestro planeta. Algunos pueden ser perjudiciales:
- Las mareas rojas causadas por algunos Dinoflagellata pueden dañar a los peces.
- Los Apicomplexa son parásitos que afectan a los animales.
- Los Oomycota causan enfermedades en las plantas. Por ejemplo, Phytophthora infestans fue responsable de una gran hambruna en Irlanda en el siglo XIX al destruir los cultivos de patata.
Sin embargo, muchos cromistas son esenciales para la vida:
- Las diatomeas son grandes productoras de oxígeno a través de la fotosíntesis. También absorben mucho dióxido de carbono, ayudando a reducir el calentamiento global.
- Las algas pardas, como el kelp, forman "bosques" submarinos donde viven muchas criaturas marinas. También son una fuente importante de alimento para las comunidades costeras.
La Simbiogénesis y la Evolución de los Cromistas

La idea de que los cromistas forman un reino se basa en la teoría de la simbiogénesis. Esta teoría explica cómo dos organismos diferentes se unen y forman uno nuevo. En el caso de los cromistas, se cree que una célula huésped (que comía otras células) se unió con un alga roja fotosintética. Con el tiempo, se fusionaron y compartieron material genético, creando una nueva entidad biológica.
Este proceso, donde una célula eucariota incorpora a otra célula eucariota fotosintética, se llama endosimbiosis secundaria. Sin embargo, los estudios genéticos han demostrado que en los cromistas no hubo un solo evento de simbiogénesis, sino varios. Esto significa que diferentes grupos de algas cromistas se originaron de forma independiente a través de distintas "capturas" de algas.
Grupos de Cromistas Fotosintéticos
Cada grupo de algas cromistas se originó por un evento de simbiogénesis particular. Aquí te mostramos algunos ejemplos:
| célula hospedadora heterótrofa + alga endosimbionte → alga resultante (nueva entidad biológica) |
|---|
| Célula huésped | Alga endosimbionte | Alga resultante | Ref. |
|---|---|---|---|
| Cryptista | Rhodophytina | Cryptophyta | |
| Heterokonta | Cryptophyceae | Ochrophyta | |
| Haptista | Ochrophyta | Haptophyta | |
| Dinoflagellata | Haptophyta | Dinophyceae | |
| Cercozoa | Chlorophytina | Chlorarachniophyceae | |
| Apicomplexa s. l. | Limnista | Chromerida |
Esto significa que algunos grupos, como las criptofíceas, se formaron por endosimbiosis secundaria. Otros, como las dinofíceas, por endosimbiosis más complejas, incluso quinarias (cinco pasos).
A veces, la unión no es completa. El organismo "capturado" mantiene algunas de sus partes, como su núcleo o mitocondrias. A esto se le llama endocitobiosis.
Ejemplos de Simbiogénesis en Cromistas
- En dinoflagelados: Algunos dinoflagelados han "robado" cloroplastos de otras algas para hacer fotosíntesis. Por ejemplo, Dinophysis se alimenta de ciliados que a su vez han capturado cloroplastos de criptofitas.
- En ciliados: Ciertas especies de ciliados marinos o de agua dulce ingieren microalgas y las mantienen vivas dentro de sí para aprovechar su fotosíntesis.
- En cercozoos: Paulinella chromatophora es un cercozoo muy especial que adquirió una cianobacteria como endosimbionte. Es un caso raro de endosimbiosis primaria en este grupo.
- En radiolarios: Muchos radiolarios tienen microalgas endosimbiontes. Estas algas les ayudan a obtener energía y son importantes para el ciclo de nutrientes en el océano.
- En foraminíferos: Los foraminíferos más grandes también albergan algas endosimbiontes. Necesitan la luz del sol para vivir, ya que dependen de sus algas.
- En criptistas: Hatena es un katablefárida que lleva un alga verde endosimbionte. Cuando se divide, solo una célula hija mantiene el alga y su capacidad fotosintética.
- En haptistas: Algunas especies de heliozoos pueden "robar" los plastos de microalgas para usarlos temporalmente.
Pérdida de la Fotosíntesis
.jpg)
A lo largo de la evolución, algunos cromistas han perdido la capacidad de hacer fotosíntesis e incluso han perdido sus plastos:
- En Pseudohongos: Los pseudohongos tienen paredes celulares con celulosa, como las algas. Aunque no tienen plastos, se han encontrado genes que sugieren que sus ancestros sí los tuvieron.
- En Apicomplexa: Los apicomplejos son parásitos que tienen un orgánulo llamado apicoplasto, que es un plasto modificado. En algunos, como Cryptosporidium, el plasto se ha perdido por completo.
- En Criptofitas: Chilomonas es una criptofita que ya no hace fotosíntesis, pero aún conserva un plasto no fotosintético.
Clasificación de los Cromistas
Según el científico Cavalier-Smith (2015), la clasificación de los cromistas puede ser la siguiente:
- reino Chromista
- subreino Hacrobia
- filo Cryptista
- subfilo Corbihelia
- subfilo Palpitia
- subfilo Rollomanidia (Chryptophyta)
- filo Haptista
- subfilo Centrohelea
- subfilo Haptophytina
- filo Cryptista
- subreino Harosa (SAR)
- infrarreino Halvaria
- superfilo Alveolata
- infrarreino Halvaria
- subreino Hacrobia
* filo Ciliophora * filo Miozoa
-
-
-
- superfilo Heterokonta (Stramenopiles)
-
-
* filo Bigyra * filo Ochrophyta * filo Pseudofungi
-
-
- infrarreino Rhizaria
- filo Cercozoa
- infrarreino Rhizaria
-
* subfilo Endomyxa * subfilo Filosa
-
-
-
- filo Retaria
-
-
* subfilo Foraminifera * subfilo Radiozoa
Galería de imágenes
-
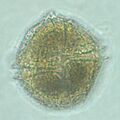
Lingulodinium, un alga Dinophyceae.
-

P. chromatophora.
-

Acantharea presentando (9) zooxantelas (fotosimbiontes) en el ectoplasma (12).



Véase también
 En inglés: Chromalveolate Facts for Kids
En inglés: Chromalveolate Facts for Kids
