
Torosaurus para niños
Datos para niños Torosaurus |
||
|---|---|---|
| Rango temporal: 72 Ma - 66 Ma Cretácico Superior | ||
 Restauración
|
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Ornithischia | |
| Suborden: | Neornithischia | |
| Infraorden: | Ceratopsia | |
| Familia: | Ceratopsidae | |
| Subfamilia: | Chasmosaurinae | |
| Género: | Torosaurus Marsh, 1891 |
|
| Especie tipo | ||
| Torosaurus latus Marsh, 1891 |
||
| Otras Especies | ||
|
||
El Torosaurus (que significa "lagarto perforado" en latín) es un género de dinosaurios ceratopsianos. Estos dinosaurios vivieron al final del período Cretácico, hace unos 72 a 66 millones de años. Sus fósiles se han encontrado en Norteamérica, desde Saskatchewan en Canadá hasta el sur de Texas en Estados Unidos.
El Torosaurus tenía uno de los cráneos más grandes de cualquier animal terrestre conocido. Su cráneo, incluyendo la gran placa ósea trasera (llamada gola o volante), podía medir hasta 2,8 metros de largo. Se estima que el Torosaurus medía alrededor de 8,8 metros de largo, 2 metros de alto y pesaba entre 4 y 6 toneladas.
Este dinosaurio se parecía mucho al Triceratops, que vivió en la misma época. Sin embargo, el Torosaurus se distinguía por su volante alargado con grandes aberturas, llamadas fenestras. También tenía huesos escamosos largos en el volante y cinco o más pares de pequeños cuernos en la parte trasera. Tenía un cuerno corto sobre su pico y dos cuernos más largos sobre sus ojos, que le servían para protegerse.
El Torosaurus: Un Gigante con Cuernos
El Torosaurus es famoso por su enorme cráneo y su distintivo volante. Este volante era una gran placa de hueso que se extendía desde la parte trasera de su cabeza.
¿Cómo era el Torosaurus?

Los Torosaurus eran dinosaurios muy grandes, similares en tamaño a los Triceratops. Su cráneo era especialmente largo debido a su volante. Se pensaba que el Torosaurus tenía el cráneo más largo de todos los animales terrestres. Sin embargo, más tarde se descubrió que el Pentaceratops podría haber tenido un cráneo aún más grande.
Características del Volante y Cuernos
El volante del Torosaurus era muy largo en comparación con el resto de su cráneo. Tenía diez o más pequeños huesos triangulares llamados epiparietales en el borde trasero. El hueso parietal (parte del volante) era delgado y tenía aberturas circulares u ovaladas.
Los cuernos sobre los ojos del Torosaurus variaban. Algunos eran grandes y curvados hacia adelante, mientras que otros eran cortos y rectos. La forma del cuerno de la nariz también cambiaba entre los individuos. Algunos tenían un cuerno nasal recto y erguido, mientras que otros solo tenían una protuberancia baja.
El volante también mostraba diferencias. Algunos volantes se curvaban hacia arriba en la parte trasera, mientras que otros eran casi planos. El número de epiparietales también variaba, aunque es difícil de saber con exactitud en muchos fósiles.
Descubrimiento y Estudios

El Torosaurus fue descubierto en 1891 en Wyoming, Estados Unidos. Fue encontrado por John Bell Hatcher, y el paleontólogo Othniel Charles Marsh le dio el nombre de Torosaurus. Se han encontrado fósiles similares en otros lugares de Norteamérica, como Montana, Dakota del Sur, Utah y Saskatchewan.
El Significado del Nombre
Aunque su nombre suena como "lagarto toro", Torosaurus en realidad significa "lagarto perforado". Esto viene de la palabra griega toreo, que significa "perforado". El nombre se refiere a los agujeros o "ventanas" en su volante, que lo distinguen del Triceratops, cuyo volante es sólido.
Especies de Torosaurus
Se han identificado dos especies principales de Torosaurus:
- Torosaurus latus: Descrita por Marsh en 1891, es la especie principal.
- Torosaurus utahensis: Descrita por Lawson en 1976.
Una tercera especie, Torosaurus gladius, se considera ahora la misma que T. latus. Los fósiles de Torosaurus son menos comunes que los de Triceratops.
¿Era el Torosaurus un Triceratops Adulto?
Ha habido un debate entre los científicos sobre si el Torosaurus y el Triceratops eran en realidad el mismo tipo de dinosaurio. Algunos investigadores, como John Scannella y Jack Horner, sugirieron en 2010 que el Torosaurus podría ser simplemente la forma adulta y madura del Triceratops.
La Hipótesis del "Toromorfo"

Esta idea se basa en que los huesos del volante de los ceratopsianos pueden cambiar mucho durante el crecimiento. Propusieron que, a medida que un Triceratops envejecía, su volante se alargaría y se harían agujeros en él, transformándose en lo que conocemos como Torosaurus. También notaron que muchos cráneos jóvenes de Triceratops tenían áreas delgadas en el volante donde podrían haberse formado los agujeros.
Argumentos en Contra

Sin embargo, otros científicos, como Andrew Farke y Nicholas Longrich, han cuestionado esta hipótesis. Argumentan que hay diferencias importantes entre el Torosaurus y el Triceratops que no se explican solo por el crecimiento. Por ejemplo, el número de pequeños huesos en el borde del volante (epoccipitales) no parece aumentar con la edad en otros ceratópsidos. Además, se han encontrado algunos fósiles de Torosaurus que parecen ser jóvenes, lo que sugiere que era un género distinto.
Los estudios más recientes, incluyendo análisis estadísticos de la forma de los cráneos, sugieren que el Torosaurus y el Triceratops eran géneros separados. Aunque el debate continúa, la mayoría de los expertos creen que son especies diferentes.
Clasificación del Torosaurus

El Torosaurus pertenece a la familia Ceratopsidae, que incluye a los dinosaurios con cuernos y picos de loro. Dentro de esta familia, se clasifica en la subfamilia Chasmosaurinae. Los análisis científicos muestran que el Torosaurus está muy relacionado con el Triceratops, siendo parientes cercanos.
Árbol Familiar de los Ceratópsidos
Los científicos usan un "árbol familiar" llamado cladograma para mostrar cómo se relacionan los dinosaurios. El Torosaurus se encuentra en una rama que incluye a otros dinosaurios con cuernos como el Anchiceratops y el Arrhinoceratops, y está muy cerca del Triceratops.
| Ceratopsidae |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galería de imágenes
-

Esqueleto montado, MPM VP6841, Milwaukee.
-
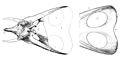
Ilustraciones originales de Marsh de los cráneos de T. latus y su sinónimo T. gladius.
-

El espécimen YPM 1831 (A) y ANSP 15192 (B).
-

Scanella & Horner vieron a Nedoceratops como una forma de transición ontogenética entre Triceratops y Torosaurus.
-

Análisis de Componentes Principales y regresión lineal entre forma y tamaño realizados en cráneos (A) y escamosos (B).
-

Espécimen MOR 1122.
.jpg)



Véase también
 En inglés: Torosaurus Facts for Kids
En inglés: Torosaurus Facts for Kids
- Anexo: Géneros válidos de dinosaurios
