
Estoma para niños


En botánica, los estomas son pequeñas aberturas que se encuentran en la epidermis de las plantas, especialmente en las hojas. La epidermis es la capa exterior de la planta, que suele ser impermeable al agua y a los gases gracias a una capa llamada cutícula. Los estomas permiten que la planta se comunique con el aire exterior.
Cada estoma está formado por dos células especiales llamadas células oclusivas, que rodean un poro central conocido como ostíolo. A veces, estas células oclusivas están acompañadas por otras células de la epidermis. Juntas, forman una estructura llamada aparato estomático. Debajo de cada estoma, hay un espacio llamado cámara subestomática, que sirve para almacenar gases. Las células oclusivas pueden abrir y cerrar el ostíolo según las necesidades de la planta, lo que es una característica importante de las plantas terrestres más evolucionadas.
Contenido
¿Para qué sirven los estomas?
Los estomas se encuentran en las partes verdes de las plantas vasculares y algunos musgos, que son la mayoría de las plantas terrestres. Estas partes verdes están cubiertas por la epidermis. Los estomas son como pequeñas puertas por donde la planta intercambia gases con el ambiente.
A través de los estomas, entra la mayor parte del dióxido de carbono y el oxígeno, gases esenciales para la fotosíntesis (cuando la planta produce su alimento) y la respiración celular. También son la vía principal por donde la planta pierde agua en forma de vapor, un proceso llamado transpiración vegetal.
Para que la planta no pierda demasiada agua, los estomas suelen estar en la parte de abajo de las hojas, donde están más protegidos del sol directo. Además, las plantas han desarrollado la capacidad de abrir o cerrar los estomas según las condiciones del ambiente y sus propias necesidades.
Intercambio de gases y agua
El dióxido de carbono es muy importante para la fotosíntesis. Las plantas necesitan que sus estomas estén abiertos durante el día para captarlo. Sin embargo, cuando los estomas están abiertos, el vapor de agua que está dentro de la hoja también se escapa. Esto significa que las plantas no pueden obtener dióxido de carbono sin perder agua al mismo tiempo.
Estrategias para el dióxido de carbono
Normalmente, el dióxido de carbono se usa en las células de la hoja para la fotosíntesis. Una enzima llamada RuBisCO es clave en este proceso. Sin embargo, RuBisCO no es muy eficiente con el dióxido de carbono y a veces usa oxígeno en su lugar, lo que hace que la planta gaste energía sin producir alimento. Por eso, RuBisCO necesita altas concentraciones de dióxido de carbono, lo que implica que los estomas deben estar muy abiertos, causando una gran pérdida de agua.
Algunas plantas usan otra estrategia. Pueden tener los estomas menos abiertos y usar una enzima diferente, la PEPcasa, que es muy buena captando dióxido de carbono. Sin embargo, usar PEPcasa requiere más energía. Esta opción es mejor cuando hay poca agua, pero mucha luz, o cuando las altas temperaturas hacen que RuBisCO funcione peor.
Plantas CAM: una adaptación especial
Hay un grupo de plantas, muchas de ellas de desiertos, llamadas plantas CAM (por "metabolismo ácido de las crasuláceas"). Estas plantas tienen una forma especial de manejar sus estomas:
- Abren sus estomas por la noche, cuando el aire es más fresco y se pierde menos agua.
- Usan la enzima PEPcasa para captar dióxido de carbono y lo guardan en grandes compartimentos dentro de sus células.
- Durante el día, cierran sus estomas y liberan el dióxido de carbono que guardaron la noche anterior para que la enzima RuBisCO lo use.
Esta estrategia les permite ahorrar mucha agua, pero tienen un límite en la cantidad de dióxido de carbono que pueden almacenar. Por eso, es una adaptación ideal para lugares con muy poca agua.
¿Cómo son los estomas?
Al observar los estomas con un microscopio, se ve que dos células, llamadas células oclusivas, son las que cambian de tamaño para regular la abertura del poro. Estas células se unen en sus extremos y forman el ostíolo.
Cuando la planta necesita cerrar el poro, las células oclusivas pierden su rigidez y se acercan, cerrando el ostíolo. Cuando necesitan abrirlo, las células se hinchan (aumentan su turgencia) y se curvan, abriendo el poro. La forma en que se curvan depende de la estructura de sus paredes celulares.
Las células oclusivas controlan su turgencia regulando la cantidad de sales de potasio (K+) dentro y fuera de ellas. El agua se mueve por ósmosis (buscando la mayor concentración de solutos) a través de la membrana celular. Este movimiento de sales y otros iones es un proceso activo que requiere energía. A veces, otras células cercanas, llamadas células acompañantes, ayudan en esta regulación.
Todas las plantas terrestres tienen una capa cerosa llamada cutícula que las cubre y las hace casi impermeables, evitando que pierdan mucha agua. Esta capa es más gruesa en plantas de lugares secos y más delgada en lugares húmedos. La cutícula solo se interrumpe en los ostíolos de los estomas. Esta capa fue crucial para que las plantas pudieran vivir en tierra, ya que las protege de la sequedad. Sin embargo, también impide el intercambio de gases, por lo que los estomas evolucionaron para permitir este intercambio vital.
Los estomas son esenciales para la fotosíntesis y la respiración de las plantas. A través de ellos, el oxígeno sale y el dióxido de carbono entra durante la fotosíntesis, y viceversa durante la respiración, según lo que la planta necesite en cada momento.
Criptas estomáticas
Las criptas estomáticas son pequeñas áreas hundidas en la epidermis de la hoja. Forman una especie de cámara que contiene uno o más estomas. A veces, también tienen pequeños pelos o cera. Estas criptas pueden ser una adaptación para ayudar a las plantas a sobrevivir en climas secos, ya que reducen la pérdida de agua. Algunas plantas que tienen criptas estomáticas son el Nerium oleander y algunas coníferas.
Estomas y la salud de las plantas
Los estomas son aberturas en la hoja por donde podrían entrar organismos que causan enfermedades (patógenos). Sin embargo, los estomas pueden detectar la presencia de algunos de estos patógenos. Curiosamente, algunas bacterias patógenas pueden producir una sustancia que hace que los estomas se vuelvan a abrir, facilitando su entrada.
Funcionamiento de los estomas
La apertura o cierre de los estomas es una parte importante de la fisiología de la planta que le permite adaptarse a las condiciones del ambiente.
Factores como la luz, la cantidad de dióxido de carbono en el aire y la disponibilidad de agua regulan muy bien la apertura y el cierre de los estomas. Se sabe que algunos iones, como el potasio, el calcio y el cloruro, participan activamente en este proceso.
Cuando hay sequía (falta de agua), los estomas se cierran para evitar que la planta pierda agua. Sin embargo, esto también impide que entre dióxido de carbono, que es necesario para la nutrición de las plantas a través de la fotosíntesis. Por eso, en regiones secas, los estomas suelen ser pequeños o casi no existen, y a menudo tienen cera o pelos que dificultan la salida del vapor de agua.
El tamaño de los estomas varía, pero generalmente miden entre 0.6 y 3.5 micrómetros (un micrómetro es una millonésima parte de un metro).
¿Cómo se forman los estomas?
Los estomas empiezan a formarse cuando una célula "madre" especial, llamada meristemática, se divide de forma desigual. Esto crea una célula pequeña (meristemoide) y una célula hermana más grande. El meristemoide puede seguir dividiéndose o convertirse en una célula madre de guarda. Finalmente, esta célula madre de guarda se divide en dos células de guarda maduras que forman el poro estomático.
Las células estomáticas tienen una pared interna más gruesa y elástica que la externa. Esta característica es muy importante para que puedan abrirse y cerrarse.
Tipos de estomas
Los científicos que estudian las plantas han observado que los estomas tienen diferentes formas y disposiciones de células. Estas diferencias son útiles para clasificar las plantas y entender sus relaciones de parentesco. Aquí te mostramos algunos tipos comunes:
- Anomocítico o ranunculáceo: No tiene células acompañantes. Es el más común en las dicotiledóneas y se considera el tipo más antiguo. Se encuentra en plantas como las de la familia Ranunculaceae.
- Paracítico o rubiáceo: Tiene 2 células acompañantes que están paralelas a las células oclusivas. Se encuentra en la familia Rubiaceae.
- Anisocítico o crucífero: Tiene 3 células acompañantes, una de ellas más pequeña. Se encuentra en las familias Cruciferae (Brassicaceae) y Solanaceae.
- Tetracítico: Tiene 4 células acompañantes. Es común en varias familias de monocotiledóneas, como Araceae, Commelinaceae y Musaceae.
- Diacítico o cariofiláceo: Tiene 2 células acompañantes que están perpendiculares a las células oclusivas. Se encuentra en pocas familias, como Cariofiláceas y Acantáceas.
- Ciclocítico: Tiene muchas células acompañantes dispuestas en uno o dos círculos alrededor de las células oclusivas.
- Helicocítico: Tiene varias células acompañantes dispuestas en espiral alrededor de las células oclusivas.
En los helechos, se distinguen cuatro tipos diferentes de estomas:
- Los estomas hipocíticos tienen dos células protectoras y dos células subsidiarias en la superficie exterior, con un poro superpuesto al estoma.
- Los estomas pericíticos tienen dos células protectoras completamente rodeadas por una célula subsidiaria continua.
- Los estomas desmocíticos tienen dos células protectoras completamente rodeadas por una célula subsidiaria que no ha unido sus extremos.
- Los estomas polocíticos tienen dos células protectoras rodeadas en gran parte por una célula subsidiaria, pero también en contacto con otras células de la epidermis.
¿Cómo evolucionaron los estomas?

No hay muchos fósiles que muestren la evolución de los estomas, pero se sabe que ya existían en las plantas terrestres a mediados del período Silúrico. Es posible que hayan evolucionado a partir de estructuras similares en los ancestros de las plantas, que eran parecidas a las algas. La evolución de los estomas debió ocurrir al mismo tiempo que la cutícula cerosa. Estos dos rasgos juntos fueron una gran ventaja para las primeras plantas que empezaron a vivir en la tierra.
Galería de imágenes
-
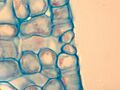
Un corte transversal de una hoja a la altura del estoma muestra la disposición de las células por debajo de él, formando una cámara subestomática que aloja los gases.
-
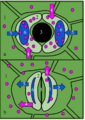
Las células oclusivas alrededor del ostíolo cambian su turgencia por ósmosis, abriéndolo o cerrándolo. El agua (flechas celestes) entra o sale de la célula oclusiva siguiendo la concentración de potasio (flechas y puntos rosas), que a su vez, entra o sale mediante estructuras en la membrana de la célula, con gasto de energía, que en conjunto se llama transporte activo.
-
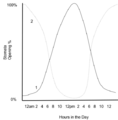
Los estomas de las plantas C3 y C4(1) permanecen abiertos todo el día y cierran por la noche. Los estomas de las plantas CAM(2) se abren durante la mañana y se cierran ligeramente al mediodía y luego se abren nuevamente por la noche.
-
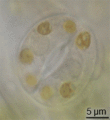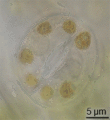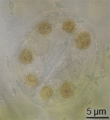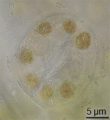
Estoma de tomate observado a través de aceite, en inmersión.
.svg)
Véase también
 En inglés: Stomata Facts for Kids
En inglés: Stomata Facts for Kids
- Lenticela
- Tejido epidérmico
- Fotosíntesis
- Nervadura
- Xilema
- Floema
