
Leishmania para niños
Datos para niños Leishmania |
||
|---|---|---|
 L. donovanii
|
||
| Taxonomía | ||
| Reino: | Protista | |
| Filo: | Euglenozoa | |
| Clase: | Kinetoplastea | |
| Orden: | Trypanosomatida | |
| Familia: | Trypanosomatidae | |
| Género: | Leishmania | |
| Especies | ||
|
Leishmania aethiopica |
||
Leishmania es un tipo de protozoo, que son organismos muy pequeños formados por una sola célula. Este protozoo es el causante de una enfermedad llamada leishmaniasis. Los principales portadores de Leishmania son unos mosquitos especiales, como los del género Phlebotomus en Europa, Asia y África, y los del género Lutzomyia en América.
Estos pequeños parásitos afectan a animales como marsupiales, cánidos (perros), roedores y primates. Se calcula que actualmente hay millones de personas en el mundo que padecen leishmaniasis.
Contenido
- Leishmania: Pequeños Parásitos con Gran Impacto
- Galería de imágenes
- Véase también
Leishmania: Pequeños Parásitos con Gran Impacto
¿Qué es Leishmania?
Leishmania es un género de protozoos que viven dentro de otros seres vivos, causando la leishmaniasis. Son tan pequeños que solo se pueden ver con un microscopio. Para pasar de un animal a otro, o de un animal a una persona, necesitan la ayuda de ciertos mosquitos.
Un Viaje en el Tiempo: La Historia de Leishmania
El origen exacto de Leishmania no está del todo claro. Algunas teorías sugieren que pudo haber surgido en África y luego se extendió a América hace millones de años, quizás a través del estrecho de Bering. Otras teorías apuntan a un origen en el paleártico (una gran región que incluye Europa y Asia).
La primera vez que se describió una enfermedad similar a la leishmaniasis fue alrededor del año 1500 d.C. en Irak. Más tarde, en 1898, un médico ruso llamado Piotr F. Borovsky fue el primero en describir el parásito en las lesiones de un paciente, pero su trabajo no fue muy conocido en ese momento.
En 1901, William Boog Leishman, un médico, observó unas pequeñas formas ovaladas en muestras de bazo de un paciente que había fallecido por una forma grave de leishmaniasis, conocida como "kala azar". Publicó sus hallazgos en 1903. Poco después, Charles Donovan, otro médico, encontró las mismas formas en otros pacientes. A estas formas se les llamó "cuerpos de Leishman-Donovan". Ellos identificaron a este protozoo como la causa del "kala azar", y lo nombraron Leishmania donovani.
Las Formas de Leishmania: Promastigote y Amastigote
Los parásitos Leishmania tienen dos formas principales durante su vida, dependiendo de dónde se encuentren:
- Promastigote: Es una forma alargada que tiene una especie de cola llamada flagelo en un extremo. Se mueve libremente y se multiplica en el intestino de los mosquitos que lo transmiten.
- Amastigote: Es una forma más redonda y pequeña. Su flagelo es tan corto que no sobresale y solo se puede ver con microscopios muy potentes. Esta forma vive y se multiplica dentro de ciertas células de los animales y humanos, llamadas macrófagos.
¿Cómo Cambia de Forma Leishmania?
Aunque parecen muy diferentes, la estructura básica de estas dos formas es similar. Ambas tienen un kinetoplasto, que es una parte especial de su mitocondria (la "central de energía" de la célula) donde guardan su ADN. El kinetoplasto está conectado al flagelo.
En el mosquito, el parásito pasa por varias etapas de promastigote. Por ejemplo, cuando el mosquito pica a un animal infectado, ingiere los amastigotes. Dentro del mosquito, estos amastigotes se transforman en promastigotes. Luego, se adhieren a las paredes del intestino del mosquito para no ser expulsados. Finalmente, se transforman en una forma especial de promastigote, llamada promastigote metacíclico, que es la forma que puede infectar a un nuevo animal o persona cuando el mosquito vuelve a picar.
El Ciclo de Vida de Leishmania: Un Viaje entre Insectos y Animales
El ciclo de vida de Leishmania es un viaje fascinante entre dos tipos de "huéspedes": un mosquito y un vertebrado (como un perro o un humano).
¿Cómo se Transmite la Leishmaniasis?
1. Una persona o animal se infecta cuando un mosquito hembra infectado lo pica. Al picar, el mosquito inyecta los promastigotes (la forma alargada del parásito) en la piel. 2. Una vez dentro del cuerpo, unas células de defensa llamadas macrófagos "se comen" a los promastigotes. 3. Dentro de los macrófagos, los promastigotes se transforman en amastigotes (la forma redonda). 4. Los amastigotes se multiplican dentro de los macrófagos y se extienden a diferentes tejidos del cuerpo. 5. Cuando un mosquito no infectado pica a una persona o animal que tiene la enfermedad, ingiere sangre con macrófagos llenos de amastigotes. Así, el mosquito se infecta. 6. Dentro del intestino del mosquito, los amastigotes se transforman de nuevo en promastigotes. 7. Estos promastigotes se multiplican en el intestino del mosquito y viajan hasta su boca. Cuando el mosquito pica a otra persona o animal, inyecta estos promastigotes, y el ciclo comienza de nuevo.
¿Dónde se Encuentra la Leishmaniasis?
La leishmaniasis se presenta de diferentes maneras, afectando la piel, las mucosas (como la nariz o la boca) o los órganos internos.
Tipos de Leishmaniasis
- Infecciones cutáneas: Son las que afectan la piel. La más conocida en el "Viejo Mundo" (Europa, Asia, África) es el "botón de Oriente", causada por especies como L. major o L. tropica. En el "Nuevo Mundo" (América), las más comunes son L. mexicana y L. braziliensis. Estas infecciones son frecuentes en países como Afganistán, Brasil, Irán, Perú, Arabia Saudita y Siria.
- Infecciones mucocutáneas: Comienzan en la piel, pero luego se extienden a las membranas mucosas, como las de la nariz, la boca o la garganta. Pueden ser graves. Son comunes en Bolivia, Brasil y Perú.
- Infecciones viscerales: Afectan los órganos internos y se caracterizan por fiebre, aumento del tamaño del hígado y el bazo, y anemia. Se conoce como "Kala azar" y es causada por el grupo L. donovani (que incluye L. donovani, L. infantum y L. chagasi). Se encuentra en zonas tropicales y subtropicales de todos los continentes, excepto Oceanía, especialmente en Bangladés, Brasil, India, Nepal y Sudán.
Galería de imágenes
-
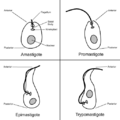
Formas celulares de los Trypanosomatida
-
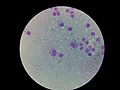
Amastigotes de Leishmania infantum, en macrófagos.
-
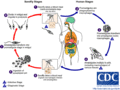
Ciclo vital de Leishmania.
-

Organización génica en Leishmania.
-
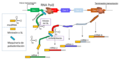
Modelo sobre la transcripción policistrónica en Leishmania, y el procesamiento mediante el mecanismo de trans-splicing (adición de un pequeño exón, denominado miniexón o SL) y la poliadenilación, para dar lugar a mensajeros individuales maduros
-
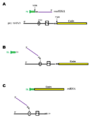
Mecanismo molecular del trans-splicing. 5’DS, sitio donador de splicing; 3’AS, sitio aceptor de splicing; A, adenosina que actúa de punto de ramificación; Py, secuencia rica en pirimidinas.





Véase también
 En inglés: Leishmania Facts for Kids
En inglés: Leishmania Facts for Kids
