
Dromiceiomimus para niños
Datos para niños Dromiceiomimus |
||
|---|---|---|
| Rango temporal: 80 Ma - 66 Ma Cretácico Superior | ||
 |
||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Ornithomimosauria | |
| Familia: | Ornithomimidae | |
| Género: | Dromiceiomimus Russell, 1972 |
|
| Especie tipo | ||
| Dromiceiomimus brevitertius Parks, 1926 |
||
| Otras Especies | ||
|
||
El Dromiceiomimus (que significa "imitador de emú" en griego) es un tipo de dinosaurio terópodo que vivió al final del periodo Cretácico. Existió hace unos 80 a 66 millones de años en lo que hoy es Norteamérica.
Este dinosaurio era un corredor muy rápido que caminaba sobre dos patas, parecido a las aves corredoras de hoy en día, como el emú. Se han encontrado esqueletos bastante completos de Dromiceiomimus, lo que ha permitido a los científicos entender bien cómo era.
Contenido
¿Cómo era el Dromiceiomimus?
El Dromiceiomimus medía aproximadamente 3.5 metros de largo y pesaba entre 100 y 150 kilogramos. Tenía patas traseras largas, ojos grandes y un pico sin dientes.
Se cree que era de sangre caliente y que probablemente tenía plumas para mantenerse abrigado. Era un animal ágil, capaz de saltar y escapar de depredadores más grandes.
Características físicas
El Dromiceiomimus tenía un cuerpo ligero y una cola corta y recta que le ayudaba a mantener el equilibrio. Su cuello era bastante largo y terminaba en una cabeza pequeña y alargada.
En lugar de dientes, tenía un pico afilado que usaba para arrancar trozos de plantas. Sus brazos eran muy largos, con tres dedos delgados y garras rectas. Estas manos le servían como ganchos para acercar ramas y alcanzar frutas o hojas.
También se piensa que podía comer pequeños animales si tenía la oportunidad, lo que lo convierte en un omnívoro. Sus patas traseras eran muy musculosas, lo que lo hacía uno de los dinosaurios más rápidos, pudiendo correr hasta 70 kilómetros por hora para huir de depredadores como el Gorgosaurus.

¿Tenía plumas el Dromiceiomimus?
Desde los años noventa, los científicos sospechaban que el Dromiceiomimus tenía plumas primitivas, parecidas a pelos, para conservar el calor corporal. En 2012, se encontraron pruebas directas de esto.
Se descubrieron tres fósiles (un joven y dos adultos) con marcas de plumas. Un espécimen adulto tenía plumas largas y reales en los brazos, similares a las de las aves. El animal joven, sin embargo, solo tenía filamentos parecidos a pelos en el cuerpo y las extremidades, lo que sugiere que las plumas más grandes se desarrollaban al crecer.
En 2016, otro fósil mostró plumas más largas en la cola que en el resto del cuerpo. También se encontraron plumas en la espalda, los costados y el vientre. Curiosamente, la parte inferior de la cola y las patas no tenían plumas, lo que podría haber ayudado al dinosaurio a refrescarse en climas cálidos.
Descubrimiento e historia
El primer fósil de Dromiceiomimus se encontró en 1924 en Alberta, Canadá. Al principio, se le dio el nombre de Struthiomimus brevetertius en 1926.
No fue hasta 1972 que el científico Dale Russell le dio su propio nombre de género, Dromiceiomimus brevitertius. En ese mismo año, otra especie, Dromiceiomimus samueli, también recibió su nombre.
Russell revisó muchos fósiles de dinosaurios parecidos a avestruces y los clasificó en tres grupos: Dromiceiomimus, Ornithomimus y Struthiomimus.
¿Por qué cambió su nombre?
El nombre original, Struthiomimus brevetertius, se refería a una característica particular de su tercer hueso del pie, que era muy estrecho. Sin embargo, algunos científicos pensaron que esta característica podría ser una variación individual o incluso una enfermedad.
En 1972, Dale Russell usó un método de estudio llamado morfometría para comparar los fósiles. Este método usa medidas y proporciones para ver qué tan similares son los especímenes. Gracias a esto, Struthiomimus brevetertius fue reconocido como un género distinto y se le dio el nombre de Dromiceiomimus.
Aunque algunos científicos han debatido si Dromiceiomimus brevitertius y Ornithomimus edmontonicus son la misma especie, hoy en día se considera que Dromiceiomimus brevitertius es el nombre válido.
¿Qué comía el Dromiceiomimus?
El paleontólogo canadiense Dale Russell sugirió que estos animales eran carnívoros, alimentándose de huevos y pequeños animales que podían sacar de sus madrigueras con sus largos brazos.
Sin embargo, otros científicos creen que su cuerpo estaba adaptado para una dieta más variada. Es probable que el Dromiceiomimus fuera omnívoro, comiendo insectos, frutas, huevos, lagartos y pequeños mamíferos. Sus ojos grandes sugieren que tenía una vista muy buena y que podría haber sido un animal nocturno.
Clasificación del Dromiceiomimus
El Dromiceiomimus pertenece a la familia de los Ornithomimidae, que son dinosaurios con características parecidas a las aves. Los estudios modernos han ayudado a ubicarlo en el árbol genealógico de los dinosaurios.
Se ha sugerido que Dromiceiomimus samueli, al ser más antiguo y muy parecido, podría ser el antepasado directo de Dromiceiomimus brevitertius.
Árbol familiar de los Ornithomimosauria
| Ornithomimosauria |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galería de imágenes
-
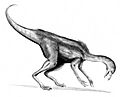
Recreación artística de Dromiceiomimus.
-

Cráneo de Dromiceiomimus.
Véase también
 En inglés: Ornithomimus Facts for Kids
En inglés: Ornithomimus Facts for Kids
