
Ácido ribonucleico para niños

El ácido ribonucleico, más conocido como ARN, es una molécula muy importante en los seres vivos. Trabaja junto con el ADN y las proteínas para que las células funcionen correctamente. El ARN se encuentra en todas las células, tanto en las más simples (procariotas) como en las más complejas (eucariotas). Algunos virus incluso usan el ARN como su único material genético.
Desde el punto de vista de la química, el ARN es como una cadena larga formada por unidades más pequeñas llamadas ribonucleótidos. Cada una de estas unidades tiene tres partes: un azúcar llamado ribosa, un fosfato y una de cuatro bases nitrogenadas: adenina (A), guanina (G), citosina (C) y uracilo (U). A diferencia del ADN, que suele tener dos cadenas, el ARN celular normalmente tiene una sola cadena. Sin embargo, algunos virus tienen ARN de doble cadena.
La función principal del ARN es ayudar a fabricar proteínas. El ADN guarda las instrucciones para hacer todas las proteínas que una célula necesita. Esta información se copia del ADN a un tipo de ARN llamado ARN mensajero (ARNm). Luego, el ARN transferente (ARNt) lleva los aminoácidos (que son los "ladrillos" de las proteínas) al ARN ribosómico (ARNr). El ARNr, que está en los ribosomas, une estos aminoácidos para construir las proteínas. El ARN también ayuda a controlar qué genes se activan o desactivan en la célula.
Contenido
- ¿Quién descubrió el ARN y cómo ha evolucionado su estudio?
- ¿Cómo está hecho el ARN?
- ¿Cómo se organiza el ARN?
- ¿Cómo se fabrica el ARN en la célula?
- ¿Qué tipos de ARN existen y para qué sirven?
- ARN en las mitocondrias
- El ARN y la información genética
- ¿Fue el ARN el origen de la vida?
- Galería de imágenes
- Véase también
¿Quién descubrió el ARN y cómo ha evolucionado su estudio?
Los ácidos nucleicos, que incluyen el ARN y el ADN, fueron descubiertos en 1867 por Friedrich Miescher. Él los llamó "nucleína" porque los encontró en el núcleo celular. Más tarde, se vio que las células sin núcleo también tenían ácidos nucleicos. En 1939, los científicos empezaron a sospechar que el ARN tenía un papel importante en la fabricación de proteínas.
Severo Ochoa ganó el Premio Nobel de Medicina en 1959 por sus descubrimientos sobre cómo se forma el ARN. En 1965, Robert W. Holley descifró la secuencia de un ARN de transferencia de una levadura, lo que le valió el Premio Nobel en 1968. En 1967, Carl Woese descubrió que algunos ARN podían actuar como catalizadores (acelerar reacciones químicas). Esto llevó a la idea de que las primeras formas de vida en la Tierra podrían haber usado ARN para guardar su información genética y para realizar sus procesos vitales.
En 1976, Walter Fiers y su equipo determinaron la secuencia completa del ARN de un virus ARN llamado MS2. En los años 90, se descubrió que el ARN podía "silenciar" genes, lo que llevó al hallazgo del ARN interferente y los micro-ARN. Estos descubrimientos han sido muy importantes para desarrollar nuevos medicamentos. En 2023, se confirmó que las moléculas de ARN fueron probablemente la primera forma de vida en nuestro planeta.
¿Cómo está hecho el ARN?


El ARN, al igual que el ADN, está formado por una cadena de unidades que se repiten, llamadas nucleótidos. Estos nucleótidos se unen entre sí mediante enlaces especiales.
Cada nucleótido tiene tres partes principales:
- Un azúcar de cinco carbonos llamado ribosa.
- Un grupo fosfato.
- Una base nitrogenada, que puede ser:
| ARN | ADN | |
|---|---|---|
| Pentosa | Ribosa | Desoxirribosa |
| Purinas | Adenina y Guanina | Adenina y Guanina |
| Pirimidinas | Citosina y Uracilo | Citosina y Timina |
Las bases nitrogenadas pueden unirse entre sí formando "pares de bases" mediante enlaces débiles. Por ejemplo, la citosina (C) se une con la guanina (G), y la adenina (A) se une con el uracilo (U).
¿Cómo se unen las bases en el ARN?

La forma más común en que las bases se unen en el ARN es similar a la del ADN: A con U (en lugar de T en ADN) y C con G. Esto se llama "apareamiento Watson y Crick". Sin embargo, en el ARN existen otras formas de unión, como el "apareamiento por balanceo" (wobble), que permite que la guanina (G) se una con el uracilo (U). Esto es importante para que el ARN de transferencia pueda leer correctamente la información.
¿Cómo se organiza el ARN?
La forma en que se organiza el ARN se describe en diferentes niveles:
Estructura primaria del ARN
La estructura primaria es simplemente la secuencia de nucleótidos en la cadena de ARN. Esta secuencia es la que contiene la información, por ejemplo, para fabricar una proteína.
Estructura secundaria del ARN

El ARN puede doblarse sobre sí mismo y formar regiones donde las bases se unen entre sí, creando "pares de bases" dentro de la misma cadena. Esto forma estructuras como tallos (regiones con bases unidas) y bucles (regiones con bases sin unir).
| Hélice
(tallo, stack) |
Región con bases apareadas. | |
|---|---|---|
| Bucle
(ciclo, loop) |
Región incluida en una hélice en donde las bases no están apareadas. |  |
| Bucle en horquilla
(tallo y bucle, hairpin loop) |
Estructura donde regiones cercanas de bases complementarias se unen, separadas por una región sin unir que permite que la secuencia se doble para formar una hélice. |  |
| Bucle interno
(internal loop) |
Estructura donde hay regiones sin unir en ambos lados de la hebra. |  |
| Protuberancia
(buldge) |
Estructura donde hay una región sin unir en un solo lado de la hebra. | |
| Bucle múltiple
(helical junction) |
Región donde se juntan varias hélices. |  |
| Pseudonudo | Una variación de bucle donde solo una parte del bucle sí está unida. |  |
_REPRESENTACI%C3%93N_3D.png)
Estructura terciaria del ARN

La estructura terciaria es la forma tridimensional final que adopta la molécula de ARN en el espacio. Esto ocurre por interacciones entre las diferentes partes de la molécula, como el apilamiento de bases o uniones especiales entre ellas.
La forma de hélice del ARN

A diferencia del ADN, que forma una doble hélice en forma de "B", las regiones de doble cadena del ARN adoptan una forma diferente, llamada "hélice A". Esta forma tiene un surco profundo y estrecho y otro más amplio y superficial. Además, el ARN es más fácil de romper que el ADN, lo que hace que las moléculas de ARN duren menos tiempo en la célula.
¿Cómo se fabrica el ARN en la célula?
La fabricación de ARN se llama transcripción. Una enzima especial, la ARN polimerasa, usa una de las cadenas del ADN como molde para crear una nueva molécula de ARN.
El proceso comienza cuando la ARN polimerasa reconoce una señal en el ADN. Luego, la enzima se mueve a lo largo de la cadena de ADN, construyendo una molécula de ARN complementaria. La secuencia del ADN también le dice a la ARN polimerasa dónde debe terminar de fabricar el ARN.
Después de la transcripción, muchos ARN son modificados por otras enzimas. Por ejemplo, al ARN mensajero de las células eucariotas se le añaden "gorros" y "colas" especiales, y se le quitan partes que no codifican proteínas (llamadas intrones).
Algunos virus también tienen sus propias enzimas que usan ARN como molde para hacer más ARN.
¿Qué tipos de ARN existen y para qué sirven?
La función principal del ARN es ayudar a fabricar proteínas, siguiendo la información del ADN. Hay varios tipos de ARN, cada uno con un papel específico:
- ARNm (ARN mensajero): Lleva la información del ADN a los ribosomas para que se fabriquen las proteínas.
- ARNt (ARN de transferencia): Transporta los aminoácidos a los ribosomas para que se unan y formen las proteínas.
- ARNr (ARN ribosómico): Forma parte de los ribosomas y ayuda a leer el ARNm y a unir los aminoácidos para construir las proteínas.
Además de estos, existen muchos otros tipos de ARN que no codifican proteínas. Se les llama ARN no codificantes y tienen funciones importantes, como regular la actividad de los genes. Algunos ARN no codificantes, llamados ribozimas, pueden incluso acelerar reacciones químicas, como cortar y unir otras moléculas de ARN o formar los enlaces entre los aminoácidos.
ARN que ayudan a fabricar proteínas
ARN mensajero (ARNm)
El ARN mensajero (ARNm) es como el "mensajero" que lleva las instrucciones del ADN (que está en el núcleo celular) a los ribosomas (que están en el citosol). En los ribosomas, estas instrucciones se usan para construir las proteínas.
ARN de transferencia (ARNt)
Los ARN de transferencia (ARNt) son moléculas pequeñas que tienen una forma especial. Su trabajo es recoger un aminoácido específico y llevarlo al ribosoma. Allí, se unen a la secuencia del ARNm para asegurarse de que el aminoácido correcto se añada a la proteína en crecimiento.
ARN ribosómico (ARNr)
El ARN ribosómico (ARNr) se combina con proteínas para formar los ribosomas, que son las "fábricas" de proteínas de la célula. El ARNr es una parte muy importante de los ribosomas y ayuda a formar los enlaces entre los aminoácidos para construir la proteína.
ARN que regulan la actividad de los genes
Muchos tipos de ARN ayudan a controlar qué genes se activan o desactivan. Lo hacen uniéndose a regiones específicas del ARNm o del ADN.
ARN de interferencia (ARNi)
Los ARN interferentes (ARNi) son moléculas pequeñas que pueden "apagar" genes específicos. Se forman a partir de moléculas de ARN más largas y actúan bloqueando la fabricación de proteínas a partir de ciertos ARNm.
Micro-ARN (miARN)
Los micro-ARN (miARN) son cadenas cortas de nucleótidos que se encuentran en las células eucariotas. Se unen a enzimas y pueden bloquear la fabricación de proteínas o hacer que el ARNm se degrade más rápido.
ARN interferente pequeño (ARNpi)
Los ARN interferentes pequeños (ARNpi o siARN) son similares a los miARN y a menudo se producen a partir de ARN de virus. Se unen a un complejo de proteínas que identifica y corta el ARNm complementario, bloqueando así la expresión del gen.
ARN asociados a Piwi
Los ARN asociados a Piwi son cadenas de nucleótidos que se encuentran en animales. Se cree que ayudan a proteger las células de elementos genéticos que se mueven (transposones) y que tienen un papel en la formación de células reproductoras.
ARN antisentido
Un ARN antisentido es una cadena de ARN que es complementaria a un ARNm. La mayoría de ellos bloquean la actividad de los genes al unirse al ARNm, formando una molécula de doble cadena que no puede usarse para fabricar proteínas y es degradada.
ARN largo no codificante (ARNlnc)
Muchos ARN largos no codificantes (ARNlnc) regulan la actividad de los genes en las células eucariotas. Un ejemplo es el Xist, que cubre uno de los dos cromosomas X en las hembras de los mamíferos, inactivándolo.
Ribointerruptor
Un interruptor molecular o ribointerruptor es una parte del ARNm que puede unirse a moléculas pequeñas. Cuando esto ocurre, afecta la actividad del gen. Así, el ARNm puede regular su propia actividad dependiendo de si la molécula señalizadora está presente o no.
ARN con actividad catalítica
Ribozimas
El ARN también puede actuar como biocatalizador, es decir, acelerar reacciones químicas. Estos ARN, llamados ribozimas, pueden realizar reacciones por sí mismos. Por ejemplo, el ARN ribosómico es una ribozima que forma los enlaces entre los aminoácidos durante la fabricación de proteínas.
Espliceosoma
Los intrones son partes del pre-ARNm que no codifican proteínas y deben ser eliminados. Esto lo hacen los espliceosomas, que contienen muchos ARN pequeños nucleares (ARNpn). En algunos casos, los propios intrones actúan como ribozimas y se eliminan a sí mismos.
ARN pequeño nucleolar (ARNpno)
Los ARN pequeños nucleolares (ARNpno) se encuentran en el nucléolo y ayudan a modificar los nucleótidos de otros ARN. Se asocian con enzimas y las guían para que realicen cambios específicos en las bases del ARN.
ARN en las mitocondrias
Las mitocondrias, que son como las "centrales de energía" de la célula, tienen su propio sistema para fabricar proteínas. Esto incluye su propio ARNr, ARNt y ARNm. Los ARN mitocondriales (ARNmt) son fabricados por una ARN polimerasa específica de las mitocondrias.
El ARN y la información genética
Aunque el ADN es la molécula que guarda la información genética en la mayoría de los organismos, el ARN también puede hacerlo.
Genomas de ARN
Los virus ARN no tienen ADN; su información genética está en el ARN. Este ARN contiene las instrucciones para fabricar las proteínas del virus y para que el virus se reproduzca.
Los "viroides" son otro tipo de patógenos que solo tienen una molécula de ARN. Este ARN no fabrica proteínas, sino que es copiado por las enzimas de la célula hospedadora (generalmente una planta).
Transcripción inversa
Algunos virus, como los retrotransposones, usan un proceso llamado transcripción inversa. Copian su ARN en ADN, y luego este ADN se usa para hacer nuevas moléculas de ARN. La telomerasa, una enzima importante en las células eucariotas, también contiene un ARN que usa como molde para construir los extremos de los cromosomas.
ARN de doble cadena

El ARN de doble cadena (dsRNA) es un tipo de ARN que tiene dos cadenas complementarias, similar al ADN, pero con uracilo en lugar de timina. Este tipo de ARN es el material genético de algunos virus. En las células eucariotas, el ARN de doble cadena puede activar el sistema de defensa del cuerpo contra las infecciones virales.
ARN circular
A finales de los años 70, se descubrió que existe una forma de ARN que es una cadena cerrada, es decir, circular. Se encuentra en animales y vegetales. Aunque su función no se conoce completamente, se ha visto que algunos ARN circulares pueden regular la actividad de los micro-ARN.
¿Fue el ARN el origen de la vida?
La hipótesis del mundo de ARN sugiere que el ARN fue la primera molécula genética en aparecer en la Tierra, antes que el ADN. Esta idea se basa en que el ARN puede guardar información genética, como el ADN, y que algunos tipos de ARN pueden realizar reacciones químicas, como las enzimas.
Durante mucho tiempo, los científicos se preguntaron qué fue primero: el ADN o las enzimas. La hipótesis del mundo de ARN resuelve este problema al proponer que las primeras formas de vida usaron el ARN tanto para guardar su información genética como para llevar a cabo sus procesos vitales.
Galería de imágenes
-
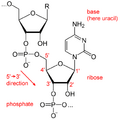
Estructura química del ARN.
-
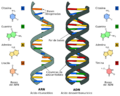
Comparación entre ARN y ADN.
-
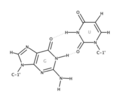
Apareamiento entre guanina y uracilo.
-

Estructura secundaria de ARN de transferencia: tRNAPhe de S. cerevisiae.
-
Bucle en la estructura secundaria del ARN.
-
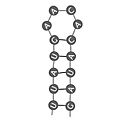
Bucle en horquilla en la estructura secundaria del ARN.
-
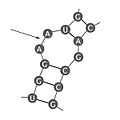
Bucle interno en la estructura secundaria del ARN.
-

Representación gráfica 3D del complejo CRISPR/Cas9
-

Bucle múltiple en la estructura secundaria del ARN.
-
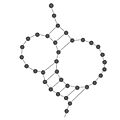
Pseudonudo en la estructura secundaria del ARN.
-
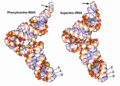
Estructura terciaria de dos ARN de transferencia: tRNAPhe y tRNAAsx.
-

Hélice A de ARN.
-
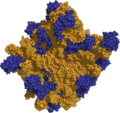
Ribosoma 50S mostrando el ARNr (amarillo), las proteínas (azul) y el centro activo, la adenina 2486 (rojo).
-

Transformación de uridina en pseudouridina, una modificación común del ARN.
-
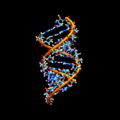
ARN bicatenario


Véase también
 En inglés: RNA Facts for Kids
En inglés: RNA Facts for Kids
