
Retrotransposón para niños
Los retrotransposones son como "genes saltarines" especiales que se encuentran en el ADN de muchos seres vivos con células complejas, como las plantas y los animales. Son un tipo de transposón que tiene la habilidad de copiarse a sí mismo y luego insertar esas copias en diferentes lugares del genoma (que es como el manual de instrucciones genéticas de un organismo).
Estos elementos son muy comunes. Por ejemplo, en el maíz, entre el 49% y el 78% de su genoma está formado por retrotransposones. En el trigo, ¡casi el 90% de su genoma son secuencias repetidas, y el 68% son elementos que pueden moverse!
En los mamíferos, casi la mitad de nuestro genoma (entre el 45% y el 48%) está compuesto por transposones o restos de ellos. En el genoma humano, alrededor del 42% son retrotransposones.
Hay dos tipos principales de retrotransposones: los LTR y los no LTR. Se clasifican según cómo son sus secuencias y cómo se mueven. En el maíz, la mayoría son LTR, mientras que en los humanos, la mayoría son no LTR. Los retrotransposones (especialmente los LTR) pueden pasar de una generación a la siguiente.
A diferencia de otros transposones que simplemente se mueven sin copiarse (lo que a veces puede causar cambios), los retrotransposones se replican. Esto significa que pueden aumentar rápidamente el tamaño del genoma y permanecer en él de forma permanente. Se cree que, al estar tanto tiempo en los genomas, desarrollaron formas especiales de insertarse que no afectan mucho el funcionamiento de los genes.
Contenido
¿Qué son los Retrotransposones LTR?


Los retrotransposones LTR (por sus siglas en inglés, que significan "repeticiones terminales largas") tienen secuencias repetidas al principio y al final, que pueden medir desde 100 hasta más de 5000 pares de bases. Estas repeticiones son características y se encuentran en cada extremo de un elemento genético que se ha integrado en el genoma de un huésped.
Los retrotransposones LTR se dividen en familias como Ty1-copia, Ty3-copia y BEL/Pao, según lo parecidas que sean sus secuencias y el orden de sus genes.
Todos los retrotransposones LTR que funcionan tienen al menos dos genes importantes: gag y pol. Estos genes son suficientes para que se puedan copiar. El gen gag produce una proteína que forma partículas parecidas a virus dentro de la célula. Dentro de estas partículas, ocurre un proceso llamado transcripción inversa. El gen pol produce otras tres proteínas: una proteasa, una transcriptasa inversa (que es una enzima clave) y una integrasa.
Generalmente, las instrucciones para los genes Gag y Pol se encuentran en el mismo ARNm (una molécula que lleva información genética). La célula huésped usa una enzima llamada ARN polimerasa II para leer estas instrucciones.
La transcripción inversa (el proceso de copiar ARN a ADN) comienza en un lugar específico. Un ARNt (otro tipo de ARN) de la célula huésped se une a este lugar y ayuda a iniciar la copia. Este proceso complejo produce una molécula de ADN de doble cadena. Finalmente, este ADN se inserta en un nuevo lugar del genoma, creando una nueva copia.
Retrotransposones Ty1-copia
Los retrotransposones Ty1-copia, también conocidos como Pseudoviridae, tienen cuatro partes importantes en sus proteínas, en este orden: proteasa, integrasa, transcriptasa inversa y ribonucleasa H.
Existen diferentes grupos dentro de los Ty1-copia, como Sirevirus/Maximus, Oryco/Ivana, Retrofit/Ale, TORK y Bianca.
Los Sirevirus/Maximus tienen un gen extra que se parece a un gen de envoltura. Este grupo fue nombrado por un elemento llamado SIRE1 encontrado en la soja (Glycine max) y también se ha visto en el maíz (Zea mays), la Arabidopsis thaliana, la remolacha (Beta vulgaris) y el pino marítimo (Pinus pinaster).
Retrotransposones Ty3-copia
Los retrotransposones Ty3-copia, o Metaviridae, tienen al menos cuatro partes importantes en sus proteínas, en este orden: proteasa, transcriptasa inversa, ribonucleasa H e integrasa. Se pueden dividir en varios grupos según su estructura y las proteínas que tienen.
Los errantivirus tienen un gen de envoltura extra que se parece al de los retrovirus. Se encontraron por primera vez en Arabidopsis thaliana y luego en otras especies como la soja y la remolacha.
Los cromovirus tienen una parte extra llamada cromodominio en su proteína integrasa. Se encuentran mucho en plantas y hongos. Se cree que este cromodominio ayuda a que el retrotransposón se inserte en lugares específicos del genoma. Los cromovirus se dividen en cuatro grupos: CRM, Tekay, Reina y Galadriel. Cada grupo se inserta en lugares distintos, como los centrómeros o los genes de ARNr.
Los elementos-ogro son retrotransposones Ty3-copia gigantes que pueden medir hasta 25.000 pares de bases. Se describieron por primera vez en el guisante (Pisum sativum).
Retrovirus endógenos
Un retrovirus endógeno es una parte de un retrovirus que se ha integrado en el genoma de un huésped y puede pasar a la siguiente generación, actuando como un retrotransposón. Comparten características con los retrovirus y los retrotransposones. Cuando el ADN retroviral se integra, se convierte en retrovirus endógenos que influyen en los genomas.
Muchos retrotransposones se parecen a los retrovirus endógenos, especialmente en su capacidad para reconocer y unirse al genoma del huésped. Sin embargo, una diferencia clave es el gen env. Si un gen es retroviral, puede haber evolucionado de un retrotransposón a un retrovirus. Los genes env se encuentran en los retrotransposones LTR de tipo Ty1-copia, Ty3-copia y BEL/Pao. Estos genes producen proteínas que ayudan a los retrovirus a entrar en las células. Los retrovirus pueden moverse entre células, mientras que los retrotransposones LTR solo pueden moverse dentro del genoma de la misma célula.
Muchos genes en los vertebrados se formaron a partir de retrovirus y retrotransposones LTR. Si un retrovirus endógeno o un retrotransposón LTR tiene la misma función y ubicación en diferentes especies, esto sugiere su importancia en la evolución.
Retrotransposones no LTR
A diferencia de los retrotransposones LTR, los retrotransposones no LTR también tienen genes para la transcriptasa inversa, proteínas que se unen al ARN y otras enzimas, pero no tienen las repeticiones largas en sus extremos. En cambio, tienen repeticiones cortas que pueden estar en orden inverso.
Aunque son retrotransposones, no pueden realizar la transcripción inversa de la misma manera que los LTR porque no tienen las secuencias que se unen al ARN.
Se dividen principalmente en dos tipos: LINE y SINE. Los elementos SVA son una excepción, ya que comparten características con ambos.
Aunque antes se les llamaba "ADN basura", las investigaciones sugieren que, en algunos casos, tanto los LINE como los SINE se han incorporado a nuevos genes para crear nuevas funciones.
Elementos nucleares largos intercalados (LINE)
Cuando un elemento LINE se copia, la copia contiene una parte que asegura que los LINE puedan copiarse en cualquier lugar donde decidan insertarse. Una enzima llamada ARN polimerasa II es la que copia los genes en ARNm. Los extremos de las copias de LINE son ricos en adeninas, lo que ayuda a que no se degraden. Esta copia es el intermediario de ARN que se mueve.
Este intermediario de ARN se mueve del núcleo al citoplasma para que se produzcan las proteínas. Luego, el ARN LINE vuelve al núcleo para insertarse en el genoma.
Los LINE se insertan en regiones del genoma que son ricas en bases A y T. Allí, usan una enzima para cortar una de las dos cadenas del ADN. La secuencia rica en adenina del LINE se une a la cadena cortada para marcar dónde se insertará. La transcriptasa inversa reconoce estas marcas para construir el retrotransposón LINE donde se cortó el ADN. Al igual que los retrotransposones LTR, este nuevo LINE insertado contiene información del genoma para que pueda copiarse y pegarse fácilmente en otras regiones.
La mayoría de las copias de LINE tienen una longitud variable al principio porque la transcripción inversa a menudo se detiene antes de terminar. Esto puede hacer que se pierda la parte que permite que se copien, impidiendo que se muevan más.
Elementos nucleares cortos intercalados (SINE)
Los SINE son mucho más cortos (unos 300 pares de bases) que los LINE. Se parecen a genes copiados por la ARN polimerasa II y la ARN polimerasa III.
Los SINE no producen su propia transcriptasa inversa y dependen de otros transposones móviles, especialmente los LINE. Los SINE usan las herramientas de los LINE, aunque las proteínas de los LINE prefieren unirse al ARN de los LINE. Los SINE no pueden moverse por sí mismos porque no pueden producir sus propias copias. Generalmente, están formados por partes derivadas de ARN y de LINE. La parte de ARNt contiene una parte que asegura que las copias de LINE se copien en ARN para que puedan seguir moviéndose. La parte de LINE permanece para que las proteínas de los LINE puedan reconocerla.
Elementos SVA
Los elementos SVA son menos comunes que los SINE y los LINE en humanos. El principio de los elementos SVA y Alu es similar, seguido de repeticiones y un final parecido al de un retrovirus endógeno. Los LINE se unen a los lados de los elementos SVA para moverlos. Los SVA son de los transposones más recientes en el genoma de los grandes simios y de los más activos en la población humana.
Un estudio reciente ha desarrollado un método para entender cómo se multiplican los elementos SVA en los genomas de los homínidos. Este método permite seguir su proliferación, identificar grupos activos y encontrar elementos "maestros" que son clave en la propagación de SVA.
Elementos Alu
Los elementos Alu son los retrotransposones más comunes en los primates. Miden unos 350 pares de bases, no producen proteínas y pueden ser reconocidos por una enzima. Su presencia puede ser importante en algunas condiciones genéticas y en el cáncer. Para que el ARN Alu se copie y pegue, necesita su extremo rico en adenina y el resto de la secuencia unida a una señal. El Alu unido a la señal puede entonces asociarse con los ribosomas. El ARN LINE también se asocia con los mismos ribosomas que el Alu. Esta unión simultánea permite que los Alu (que son SINE) interactúen con los LINE, lo que facilita que los SINE se copien y peguen.
Retroposones
Los retroposones son retrotransposones que se insertan en los cromosomas después de haber sido copiados de cualquier molécula de ARN, pero sin producir su propia transcriptasa inversa. Por lo tanto, no pueden moverse por sí mismos. A veces, los retrotransposones no LTR, como los elementos LINE1 humanos, son llamados retroposones, pero esto depende de la definición.
Los retroposones han causado unas 10.000 duplicaciones de genes en el genoma humano, y se cree que entre el 2% y el 10% de estos genes son funcionales. Estos genes se llaman retrogenes y son un tipo de retroposón. Un ejemplo clásico es la copia de una molécula de ARN de un gen llamado c-src en el ancestro de un virus.
Retrozimas
Las retrozimas, presentes en plantas y animales, son retrotransposones no LTR que no pueden moverse por sí mismos y, por lo tanto, "piden prestadas" proteínas de otros transposones para moverse a nuevas regiones del genoma. Se copian activamente en ARN circulares usando un método de "replicación en círculo rodante" y una ribozima (una molécula de ARN con actividad enzimática). La secuencia de retrozima es copiada primero por una enzima del huésped, produciendo una secuencia de ARN que contiene varias copias de la retrozima. Luego, la ribozima se corta a sí misma para separar las copias en moléculas individuales.
Papel en la evolución
Los análisis sugieren que los retrotransposones se originaron de elementos genéticos más antiguos en organismos simples, antes de que aparecieran los seres vivos con células complejas. Los retrotransposones LTR aparecieron más tarde que los no LTR, posiblemente a partir de un retrotransposón no LTR ancestral que adquirió una enzima llamada integrasa de otro tipo de transposón. Los retrovirus obtuvieron propiedades adicionales en sus envolturas al tomar genes de otros virus, usando el poder de los retrotransposones LTR.
Debido a su forma de copiarse, los retrotransposones aumentan rápidamente en número, llegando a componer el 40% del genoma humano. Las tasas de inserción de los elementos LINE1, Alu y SVA varían, y las tasas de LINE1 han cambiado mucho en los últimos 35 millones de años, lo que nos da pistas sobre la evolución del genoma.
Es interesante que una gran parte del genoma del maíz (100.000 pares de bases) muestra mucha variedad debido a la presencia o ausencia de retrotransposones. Sin embargo, como el maíz es genéticamente inusual en comparación con otras plantas, no se puede usar para predecir cómo se mueven los retrotransposones en otras plantas.
Implicación en la salud
Los cambios causados por los retrotransposones pueden incluir:
- Desactivación de genes.
- Cambios en cómo se regulan los genes.
- Modificación de los productos de los genes.
- Actuar como lugares para la reparación del ADN.
Por lo tanto, están relacionados con el desarrollo de diferentes condiciones genéticas, como por ejemplo:
- Cáncer: Los transposones pueden insertarse en genes importantes para el control del crecimiento celular, como los que suprimen tumores, y hacer que la proteína deje de funcionar. La actividad de retrotransposones como los de la familia LINE-1 se ha asociado con el desarrollo de cáncer de colon, mama, próstata o pulmón.
- Hemofilia A: Las inserciones de transposones, como LINE-1 y Alu, pueden interrumpir un gen clave, causando problemas en la coagulación de la sangre.
- Condiciones neurológicas: Como el autismo u otras condiciones que afectan el cerebro.
- Ataxia telangiectasia: Se ha visto que las inserciones de elementos Alu o de pseudogenes en un gen específico pueden causar la pérdida de función de una proteína, llevando al desarrollo de esta condición.
Clasificación
Actualmente, los transposones se clasifican de la siguiente manera y se reconocen las siguientes familias de retrotransposones:
- Retrotransposones LTR
- Ty1-copia (Pseudoviridae)
- Ty2-copia (Metaviridae)
- Bel/pao (Belpaoviridae)
- Retrovirus endógeno (Retroviridae)
- Retrotransposones no LTR
Galería de imágenes
-

Retrotransposón.
-
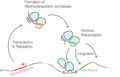
Mecanismo de retroposición.
Véase también
- Retroposón
- Transposones de ADN
- Plásmido
- Secuencia de inserción
- Retrovirus endógeno
- Virus retrotranscrito
