
Micro-ARN para niños
El micro-ARN o mi-ARN es una molécula muy pequeña de ARN, como un hilo simple, que mide entre 21 y 25 nucleótidos (los "ladrillos" que forman el ARN). Su trabajo principal es ayudar a controlar cómo funcionan otros genes en nuestras células. Lo hace a través de un proceso llamado ribointerferencia.
Los micro-ARN fueron descubiertos por primera vez en 1993 por Rosalind Lee y su equipo. Sin embargo, el nombre "micro-ARN" se usó oficialmente en 2001. A principios de 2008, los científicos estimaron que podría haber alrededor de 50,000 tipos diferentes de micro-ARN en el genoma humano, y cada uno podría influir en miles de otros micro-ARN.
Contenido
¿Qué son los micro-ARN?
Después de descubrir otras moléculas de ARN llamadas siARN, los investigadores se preguntaron si las células también tenían proteínas que ayudaban a eliminar el ARN mensajero (ARNm). Al buscar, no encontraron siARN, sino los micro-ARN, que ya se habían identificado antes.
Los micro-ARN son moléculas de ARN que se crean a partir de la información en el ADN. A diferencia de otros ARN, los micro-ARN no se usan para fabricar proteínas. Se encuentran en muchos seres vivos, desde plantas hasta gusanos y seres humanos. Muchos micro-ARN son muy parecidos entre diferentes especies, lo que sugiere que son muy antiguos en la historia de la vida. Se calcula que los micro-ARN podrían representar al menos el 3% de todos los genes humanos.
La parte del ADN que contiene la información para un micro-ARN es más larga que el micro-ARN final. Esta secuencia de ADN tiene una parte que puede doblarse y unirse consigo misma, formando una estructura con forma de horquilla. Cuando esta secuencia de ADN se copia (un proceso llamado transcripción genética), se forma un ARN primario largo conocido como pri-micro-ARN.
Luego, una enzima especial en el núcleo celular llamada DROSHA corta esta horquilla, creando lo que se llama pre-micro-ARN. Este pre-micro-ARN viaja desde el núcleo hasta el citoplasma (la parte de la célula fuera del núcleo). Una vez en el citoplasma, otra enzima llamada DICER lo corta hasta su tamaño final de 20-25 nucleótidos.
¿Cómo funcionan los micro-ARN?
La función principal de los micro-ARN es regular la actividad de los genes. Un micro-ARN se une a una parte de uno o más ARN mensajeros (ARNm). En los animales, los micro-ARN suelen unirse de forma "imperfecta" y generalmente impiden que el ARNm se convierta en proteína. En las plantas, la unión suele ser "perfecta" y hace que el ARNm se rompa y se degrade.
Al principio, los micro-ARN se descubrieron en gusanos, donde controlan las etapas del crecimiento. Hoy sabemos que están involucrados en muchos procesos de desarrollo y ayudan a ajustar finamente la actividad de los genes en la célula. Como un solo micro-ARN puede influir en miles de genes (alrededor del 30% de los genes humanos), son una parte importante de cómo las células controlan su funcionamiento. Si los micro-ARN no funcionan correctamente, pueden surgir problemas en la célula.
Características de los micro-ARN
Los micro-ARN se crean a partir de diferentes lugares en el genoma como moléculas primarias largas llamadas pri-micro-ARN.
Los micro-ARN pueden encontrarse en varias partes del genoma:
- Algunos micro-ARN tienen sus propios "interruptores" (promotores) que controlan cuándo se activan.
- La mayoría de los micro-ARN se encuentran dentro de otras partes de los genes llamadas intrones. Se copian junto con el gen principal y luego se separan.
- Otros micro-ARN se encuentran en las regiones que no se usan para fabricar proteínas (llamadas UTR 5' o 3') de otros genes.
Aproximadamente la mitad de los micro-ARN se encuentran en "grupos" (clusters) en el genoma. Estos grupos se copian juntos como una sola molécula grande que luego se divide en varios micro-ARN. Generalmente, los micro-ARN de un mismo grupo se activan al mismo tiempo.
Cómo se forman y procesan los micro-ARN

Los genes que producen micro-ARN son mucho más largos que los micro-ARN finales. Primero, se transcriben como moléculas primarias (pri-micro-ARN) y luego se procesan en el núcleo celular en estructuras más cortas con forma de horquilla, llamadas pre-micro-ARN. En animales, este procesamiento lo realiza un grupo de proteínas llamado Microprocesador, que incluye la nucleasa DROSHA y la proteína PASHA.
Estos pre-micro-ARN son luego procesados en el citoplasma por la ribonucleasa DICER, que también ayuda a formar el complejo RISC (ARN-induced silencing complex). Este complejo es el que silencia los genes. El proceso es un poco diferente en las plantas, ya que no tienen una proteína como DROSHA. También es diferente para los micro-ARN que vienen de intrones, que son procesados solo por DICER.
DROSHA es una proteína grande que trabaja junto con otra proteína llamada PASHA (o DGCR8 en mamíferos). DGCR8 ayuda a DROSHA a reconocer la parte correcta del ARN y a cortarlo a la distancia adecuada.
Para que DROSHA funcione bien, el ARN con forma de horquilla debe tener "colas" largas de ARN de una sola hebra en ambos extremos. Una vez que se forma el pre-micro-ARN, DICER lo corta y crea dos moléculas cortas. Solo una de ellas se une al complejo RISC. Esta hebra se llama "hebra guía" y es elegida por una proteína llamada Argonauta. La otra hebra se desecha.
Una vez que la hebra guía está en el complejo RISC activado, el micro-ARN se une a un ARNm complementario. En animales, esto generalmente impide que el ARNm se convierta en proteína.
Aunque DICER es muy importante, se ha descubierto que algunos micro-ARN pueden formarse sin DICER, usando la actividad de otra proteína llamada Argonauta2 (Ago2). Esto significa que hay diferentes maneras en que los micro-ARN pueden ser creados en la célula.
Las proteínas Argonauta, que son parte del complejo RISC, se encuentran en lugares especiales del citoplasma llamados "cuerpos-P". Estos cuerpos-P son lugares donde los ARNm se degradan rápidamente. Se pensaba que los cuerpos-P eran esenciales para el proceso de silenciamiento de genes, pero estudios posteriores han demostrado que las células pueden silenciar genes incluso sin ellos.
¿Cómo regulan los micro-ARN la expresión de los genes?
Aunque se ha avanzado mucho en entender cómo se forman y funcionan los micro-ARN, los científicos aún debaten los mecanismos exactos. Se ha visto que los micro-ARN en células animales pueden reducir la actividad de los genes de varias maneras:
- Pueden hacer que las proteínas se degraden durante su formación.
- Pueden detener la formación de proteínas.
- Pueden hacer que la formación de proteínas termine antes de tiempo.
- Pueden impedir que la formación de proteínas comience.
Además, los micro-ARN en animales pueden causar la degradación de los ARNm a los que se unen, incluso si la unión no es perfecta. Esto lo hacen quitando las "tapas" de los extremos del ARNm. También pueden "esconder" los ARNm en los cuerpos-P, donde no pueden ser usados para hacer proteínas. Los científicos siguen investigando para entender mejor estos mecanismos.
En algunos casos, se ha descubierto que los micro-ARN también pueden activar la producción de proteínas.
Características generales de los micro-ARN:
- Se unen a una región específica de los ARNm.
- Generalmente, se necesitan varios sitios de unión para que un micro-ARN tenga un efecto significativo.
- Un mismo ARNm puede ser regulado por diferentes micro-ARN.
- Un solo micro-ARN puede controlar la actividad de cientos de ARNm diferentes. Los estudios han mostrado que los micro-ARN suelen reducir los niveles de proteínas de forma sutil, pero estos pequeños cambios pueden tener un gran impacto en cómo funciona la célula.
- La especificidad de un micro-ARN (a qué ARNm se une) está determinada por una pequeña parte de su secuencia, llamada "región semilla".
- Un micro-ARN puede funcionar incluso si no se ha formado en el núcleo de la célula.
- El micro-ARN es el que decide a qué ARNm se unirá el complejo RISC.
Funciones biológicas de los micro-ARN
Función en la respuesta del cuerpo
- Defensa contra virus: Algunos micro-ARN pueden ayudar a las células a defenderse de los virus. Por ejemplo, el micro-ARN miR-32 puede unirse a una secuencia de ARN de ciertos virus y detener su actividad.
- Ayuda a los virus: Curiosamente, algunos micro-ARN pueden ayudar a los virus a funcionar mejor. Por ejemplo, el micro-ARN miR-122, que se encuentra en el hígado, es necesario para que el virus de la hepatitis C se active de manera eficiente.
- Muchos micro-ARN tienen papeles importantes en el desarrollo y funcionamiento de las células del sistema inmunitario, que nos defienden de enfermedades.
Función en el desarrollo
En algunos organismos, como el gusano C. elegans, los micro-ARN ayudan a que el organismo pase rápidamente por las diferentes etapas de su crecimiento. Por ejemplo, los micro-ARN lin-4 y let-7 controlan cuándo las células nerviosas y de la piel se desarrollan durante las etapas tempranas. Estos micro-ARN y sus funciones se están estudiando activamente en mamíferos.
En las células musculares de mamíferos, los micro-ARN también tienen un papel. Por ejemplo, la activación de un factor llamado SRF hace que se produzcan los micro-ARN miR-1-1 y miR-1-2, que a su vez controlan otras proteínas importantes para el desarrollo muscular.
Uno de los primeros micro-ARN descubiertos con una función en el desarrollo del sistema inmunitario fue miR-181a. Este micro-ARN se encuentra en altas cantidades en las células del timo (un órgano del sistema inmunitario) y en menores cantidades en otras células. Se ha demostrado que miR-181a puede influir en el desarrollo de diferentes tipos de células del sistema inmunitario.
También se ha visto que los micro-ARN se distribuyen de forma específica en diferentes partes del cuerpo. Por ejemplo, en los embriones de pez cebra, miR-206 se encuentra principalmente en el músculo, miR-126 en los vasos sanguíneos y el corazón, y miR-30c en los riñones. Esto sugiere que su actividad está limitada a los tejidos y órganos donde se expresan.
Nuevas funciones en el desarrollo temprano
Estudios recientes en ratones han mostrado que una cantidad importante de micro-ARN de la madre se transmiten a los embriones en las primeras etapas del desarrollo. Esto sugiere que estos micro-ARN maternos podrían tener un papel importante en el desarrollo temprano del embrión.
Control del colesterol y los triglicéridos
Los estudios han identificado ciertos micro-ARN que son importantes para controlar el colesterol, los triglicéridos y el uso de energía en el cuerpo.
Los micro-ARN como miR-128-1 y miR-148a son muy importantes en este control. Por ejemplo, si se introducen estos micro-ARN en células del hígado humano, se reduce la producción de proteínas que ayudan a eliminar el colesterol "malo" (LDL) y a producir colesterol "bueno" (HDL). Esto significa que estos micro-ARN pueden influir en los niveles de colesterol en la sangre.
Si se bloquea la acción de estos micro-ARN, la producción de las proteínas que eliminan el colesterol "malo" y producen el "bueno" aumenta. Esto sugiere que los micro-ARN podrían ser importantes para entender y tratar problemas relacionados con el colesterol y los lípidos en la sangre.
Galería de imágenes
-
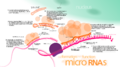
Diagrama de micro-ARN en acción con ARN mensajero
-
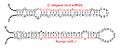
Ejemplos de bucles de micro-ARN; los micro-ARN maduros aparecen en rojo.
-
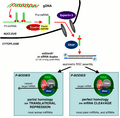
Ilustración de las principales diferencias entre el silenciamiento de genes entre plantas y animales. Los micro-ARN endógenos o los siARN son procesados por DICER e integrados en el complejo RISC, que media el silenciamiento de genes.
-

La estructura secundaria en stem-loop (tallo-asa) de un pre-micro-ARN de Brassica oleracea.



Véase también
 En inglés: MicroRNA Facts for Kids
En inglés: MicroRNA Facts for Kids
