
Reparación del ADN para niños
La reparación del ADN es un conjunto de procesos vitales que ocurren dentro de nuestras células. Imagina que el ADN es como un libro de instrucciones muy importante para tu cuerpo. Este libro contiene toda la información que tu cuerpo necesita para funcionar. A veces, este libro puede sufrir daños, ya sea por cosas que pasan dentro de la célula (como las actividades normales del cuerpo) o por factores externos (como la luz del sol o la radiación).
Estos daños pueden ocurrir hasta un millón de veces al día en cada célula. Si no se reparan, pueden causar problemas en la forma en que la célula usa su información genética. Esto podría llevar a que la célula no funcione bien o incluso a que se desarrolle un crecimiento celular descontrolado. Por eso, las células tienen un equipo especial de "reparadores" que trabajan constantemente para encontrar y arreglar estos daños en el ADN.
La rapidez con la que se repara el ADN depende de varios factores, como el tipo de célula, su edad y el ambiente que la rodea. Si una célula acumula muchos daños en su ADN o no puede repararlos bien, puede pasar por uno de estos caminos:
- Entrar en un estado de inactividad permanente, llamado senescencia.
- Activar su propia "autodestrucción", un proceso llamado apoptosis o muerte celular programada.
- Desarrollar un crecimiento celular descontrolado, que puede llevar a enfermedades graves.
La capacidad de reparar el ADN es muy importante para mantener la información genética de las células en buen estado. Esto asegura que el cuerpo funcione correctamente. Incluso se ha descubierto que muchos genes relacionados con la longevidad (cuánto tiempo vive un organismo) también tienen un papel en la reparación y protección del ADN. Si los daños no se corrigen en las células que forman las células reproductoras, pueden aparecer cambios en la información genética de los descendientes, lo que influye en la evolución.
Contenido
¿Qué es el daño al ADN?

Los daños al ADN ocurren constantemente debido a factores del ambiente y a los procesos normales que suceden dentro de la célula. Se estima que cada célula sufre entre mil y un millón de "lesiones" moleculares al día. Aunque esto parece mucho, es una parte muy pequeña del total de información genética humana. Sin embargo, si estas lesiones no se reparan en genes clave (como los que controlan el crecimiento celular), pueden impedir que una célula funcione bien y aumentar la posibilidad de un crecimiento celular descontrolado.
La mayoría de los daños al ADN afectan la estructura principal de la doble hélice, que es como una escalera retorcida. Esto significa que la química de las "bases" (los bloques de construcción del ADN) se modifica. Estas modificaciones pueden alterar la forma normal de la hélice, añadiendo nuevas conexiones químicas o "bultos" que no encajan. A diferencia de las proteínas, el ADN no suele tener una estructura tridimensional compleja, pero sí se enrolla alrededor de unas proteínas llamadas histonas (en las células eucariotas), y ambas estructuras pueden dañarse.
¿Qué causa los daños al ADN?
Los daños al ADN se pueden clasificar en dos tipos principales:
- Daños internos (endógenos): Son causados por procesos normales dentro de la célula, como el ataque de moléculas reactivas de oxígeno que se producen durante el metabolismo.
- Daños externos (exógenos): Son causados por agentes de fuera de la célula, como:
* Radiación ultravioleta (UV) del sol u otros tipos de radiación, como los rayos X y los rayos gamma. * Radiación ionizante. * Calor o Hidrólisis (reacciones con agua). * Algunas sustancias tóxicas de plantas. * Sustancias químicas artificiales que pueden alterar el ADN. * Tratamientos médicos como la Quimioterapia y la radioterapia para enfermedades graves. * Virus que se integran en la información genética de la célula.
Si el ADN dañado se copia antes de que la célula se divida, pueden incorporarse "letras" incorrectas. Las células hijas que heredan estas "letras" equivocadas tendrán cambios permanentes en su información genética.
Tipos de daños al ADN
Hay cuatro tipos principales de daños al ADN causados por procesos internos de la célula:
- Oxidación de las bases (como la 8-oxoG) y roturas en la cadena de ADN por moléculas de oxígeno reactivas.
- Adición de grupos químicos a las bases (normalmente metilación), como la formación de 7-metilguanina.
- Hidrólisis de bases, como la desaminación (pérdida de un grupo amino).
- Errores en la copia del ADN, donde se inserta una base incorrecta, se añade una base extra o falta una base.
Los daños causados por agentes externos son de muchos tipos. Algunos ejemplos son:
- La luz UV-B del sol puede hacer que bases cercanas como la citosina y la timina se unan de forma anormal, formando "dímeros de pirimidina". Esto se llama daño directo al ADN.
- La luz UV-A puede generar "radicales libres", especialmente si hay sustancias como protectores solares que han penetrado en la piel. Esto se llama daño indirecto al ADN.
- La radiación ionizante, como la de la desintegración radiactiva o los rayos cósmicos, puede romper las cadenas de ADN.
- El calor a temperaturas elevadas aumenta la pérdida de bases y las roturas de cadenas sencillas. Por ejemplo, en bacterias que viven en aguas termales muy calientes, la pérdida de bases es muy alta.
- Sustancias químicas industriales (como el cloruro de vinilo) y compuestos ambientales (como los hidrocarburos policíclicos del humo) pueden causar muchos tipos de daños al ADN, como bases oxidadas o uniones anormales.
Los daños por radiación UV, la adición de grupos químicos, los rayos X y los daños por oxidación son ejemplos de daños inducidos. Los daños espontáneos incluyen la pérdida de una base, la desaminación y errores en la estructura del azúcar.
Daños en el ADN del núcleo y de las mitocondrias
En las células humanas, el ADN se encuentra en dos lugares: en el núcleo y en las mitocondrias. El ADN nuclear (ADNn) está muy organizado y enrollado alrededor de proteínas llamadas histonas, formando los cromosomas. Cuando una célula necesita usar la información de su ADNn, la región correspondiente se desenrolla, se usa la información y luego vuelve a enrollarse.
El ADN mitocondrial (ADNmt) se encuentra dentro de las mitocondrias (que son como las "centrales de energía" de la célula). Hay muchas copias de ADNmt en cada mitocondria. Dentro de las mitocondrias, las moléculas reactivas de oxígeno y los radicales libres, que son subproductos de la producción de energía, crean un ambiente que puede dañar el ADNmt. Una enzima importante que ayuda a proteger el ADNmt es la superóxido dismutasa.
Envejecimiento y apoptosis
El envejecimiento celular es un estado en el que la célula deja de dividirse. Es una forma de protección relacionada con el acortamiento de los extremos de los cromosomas, llamados telómeros. Los telómeros son regiones repetitivas de ADN que se acortan un poco cada vez que una célula se divide. El envejecimiento de las células puede ser una alternativa a la apoptosis (autodestrucción) cuando la célula es necesaria para el organismo, pero tiene el ADN dañado y se quiere evitar que se copie de forma anormal. La división celular sin control puede llevar a un crecimiento celular descontrolado, lo que es peligroso para el organismo. Por eso, el envejecimiento y la apoptosis son parte de la estrategia de protección contra estas enfermedades.
Daños en el ADN y cambios genéticos
Es importante diferenciar entre los daños en el ADN y los cambios genéticos (llamados mutaciones). Los daños en el ADN son alteraciones químicas en su estructura, como roturas o adiciones de sustancias. Algunas proteínas pueden reconocer estos daños y repararlos si hay una copia intacta de la información en la otra cadena del ADN. Si una célula no repara los daños, la información de un gen puede dejar de usarse.
En cambio, una mutación es un cambio en la secuencia de "letras" del ADN. La estructura química de las bases es "normal", por lo que las proteínas reparadoras no pueden reconocer el cambio. Una vez que una mutación está presente en ambas cadenas del ADN, es muy difícil de corregir y se copia durante la división celular.
A nivel celular, las mutaciones pueden cambiar cómo funciona una célula. En un organismo, el número de células con mutaciones puede aumentar o disminuir según cómo afecte la mutación a la capacidad de la célula para sobrevivir y reproducirse. Aunque son diferentes, los daños en el ADN y las mutaciones están relacionados, ya que los daños a menudo causan errores al copiar o reparar el ADN, y estos errores son una causa importante de mutaciones.
Los daños en el ADN son un problema especial en células que no se dividen o lo hacen lentamente, porque los daños no reparados tienden a acumularse con el tiempo. En células que se dividen rápidamente, los daños no reparados que no matan la célula suelen causar errores durante la copia del ADN y, por lo tanto, mutaciones.
La mayoría de las mutaciones que no son neutras son perjudiciales para la supervivencia de una célula. Sin embargo, algunas mutaciones pueden dar una ventaja a la célula para crecer y dividirse, lo que puede llevar a un crecimiento celular descontrolado. Las células que se dividen más a menudo tienen más probabilidades de acumular mutaciones. Por lo tanto, los daños en el ADN en células que se dividen frecuentemente son una causa importante de enfermedades graves relacionadas con el crecimiento celular. En cambio, los daños en el ADN en células que se dividen poco son probablemente una causa importante del envejecimiento.
¿Cómo se repara el ADN?

Las células no pueden funcionar si los daños en el ADN afectan la información esencial. Dependiendo del tipo de daño, las células han desarrollado varias formas de reparar el ADN y restaurar la información perdida. Si es posible, las células usan la cadena de ADN complementaria (la que no está dañada) o una cromátida hermana (una copia idéntica del cromosoma) como "plantilla" para restaurar la información original. Si no hay una plantilla disponible, las células usan un sistema de emergencia que puede cometer errores, llamado síntesis de translesión.
Los daños al ADN cambian la forma de la hélice, y la célula puede detectar estos cambios. Una vez que se detecta el daño, unas moléculas reparadoras especiales se unen al lugar dañado, atrayendo a otras moléculas para formar un equipo que permite la reparación. Los tipos de moléculas y el mecanismo de reparación que se usan dependen del tipo de daño y de la fase del ciclo celular en que se encuentre la célula.
Daños en una sola cadena
Cuando solo una de las dos cadenas de la doble hélice está dañada, la otra cadena puede usarse como guía para corregir la parte dañada. Para reparar estos daños, existen varios mecanismos que eliminan el nucleótido dañado y lo reemplazan con uno nuevo que coincide con el de la cadena no dañada.
- Reparación sobre la marcha: Es el sistema principal de corrección de errores que cometen las propias enzimas que copian el ADN (ADN Polimerasas). Si colocan un nucleótido equivocado, lo detectan por la distorsión que causa en la doble hélice y lo corrigen inmediatamente.
- Reparación directa: No elimina nucleótidos. En cambio, usa enzimas especiales para reparar directamente las alteraciones. Por ejemplo, la fotoliasa puede separar los dímeros de timina causados por la luz UV.
- Reparación por escisión de base (BER): Repara daños a un solo nucleótido causados por oxidación, adición de grupos químicos, hidrólisis o desaminación. Una enzima quita la base dañada, luego otra enzima quita el resto del nucleótido, y finalmente se coloca el nucleótido correcto.
- Reparación por escisión de nucleótido (NER): Repara daños que afectan segmentos más largos de ADN (entre dos y treinta bases). Este proceso reconoce cambios grandes que distorsionan la hélice, como los dímeros de timina. Una forma especial de NER se enfoca en genes que se están usando activamente.
- Reparación de malapareamiento (MMR): Este sistema corrige errores de nucleótidos que se emparejaron mal (pero que no están dañados) después de que la copia del ADN ha terminado. En bacterias, este sistema sabe qué cadena es la nueva (y por lo tanto la que tiene el error) porque la cadena original está marcada. En células humanas, el mecanismo para distinguir la cadena nueva de la vieja aún se está investigando.
Estos métodos reparan el ADN de forma muy precisa, recuperando la información original. Pero cuando los daños son demasiados, se activan otros tipos de reparación que pueden cometer errores: son soluciones de emergencia para la supervivencia de la célula.
- Respuesta SOS: Es un sistema de emergencia que se activa cuando se acumulan daños que distorsionan la doble hélice y detienen la maquinaria de copia del ADN. En bacterias, esto activa enzimas que pueden "saltarse" la distorsión y rellenar los huecos con nucleótidos al azar. En células humanas, estas enzimas están siempre presentes, pero solo se activan cuando hay muchos daños.
- Proteína p53: Más que un sistema de reparación, esta proteína puede inducir la autodestrucción de la célula (apoptosis) si los daños no pueden repararse, para evitar que se desarrollen crecimientos celulares descontrolados.
Daños en doble cadena
Las roturas en doble cadena, donde ambas cadenas de la doble hélice se rompen, son muy peligrosas para la célula porque pueden causar grandes problemas en la información genética. Existen dos mecanismos para reparar estas roturas: la unión de extremos no homólogos (NHEJ) y la reparación recombinativa.
En el NHEJ, una enzima especial (ADN ligasa IV) une directamente los dos extremos rotos. Para una reparación precisa, el NHEJ busca secuencias cortas similares en los extremos del ADN que deben unirse. Si estas secuencias son compatibles, la reparación suele ser correcta. Sin embargo, el NHEJ también puede causar cambios genéticos durante la reparación, como la pérdida de bases o la unión de fragmentos incorrectos. El NHEJ es especialmente importante antes de que la célula haya copiado su ADN, ya que no hay una plantilla para la reparación recombinativa.
La reparación recombinante necesita la presencia de una secuencia idéntica o casi idéntica que se usa como plantilla para reparar la rotura. Las enzimas que realizan este proceso son muy similares a las que causan el cruce cromosómico durante la meiosis (un tipo de división celular). Esta vía permite que un cromosoma dañado se repare usando una cromátida hermana (disponible después de que el ADN se ha copiado) o un cromosoma homólogo como plantilla. Las roturas de doble cadena causadas por intentos de copiar el ADN a través de un daño suelen repararse por recombinación.
Algunas enzimas llamadas topoisomerasas causan roturas temporales en una o ambas cadenas del ADN para cambiar su forma. Estas roturas no se consideran daños, ya que son parte del proceso normal de estas enzimas y se reparan inmediatamente.
Científicos franceses estudiaron cómo la bacteria Deinococcus radiodurans repara las roturas de doble cadena. Descubrieron que esta bacteria, que es muy resistente a la radiación, puede usar múltiples copias de su información genética para reconstruir los fragmentos rotos, utilizando un proceso de recombinación homóloga muy eficiente.
Galería de imágenes
-
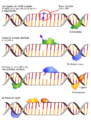
La reparación por escisión de bases es una de las formas más sencillas de reparación del ADN. en la imagen se muestra en su versión más corta.
Véase también
 En inglés: DNA repair Facts for Kids
En inglés: DNA repair Facts for Kids
