
Transducción de señal para niños
La transducción de señal es como las células de nuestro cuerpo se comunican y responden a los mensajes que reciben de su entorno. Imagina que una célula es como una casa y los mensajes son cartas que llegan a la puerta. La transducción de señal es el proceso por el cual la casa "lee" la carta y decide qué hacer.
Este proceso tiene dos pasos principales:
- Una molécula de señalización (la "carta") se une a un receptor especial en la superficie de la célula (la "puerta").
- El receptor envía la señal al interior de la célula, usando otras moléculas llamadas "segundos mensajeros", lo que provoca una respuesta.
Lo interesante es que una sola molécula de señalización puede causar muchas respuestas dentro de la célula, como si una carta pudiera activar muchas cosas a la vez.
Contenido
- ¿Cómo se comunican las células?
- ¿Qué activa a las células?
- ¿Qué hacen las células en respuesta?
- Tipos de mensajes celulares
- Tipos de receptores celulares
- Amplificación de la señal
- Transducción de señal dentro de la célula
- Galería de imágenes
- Véase también
¿Cómo se comunican las células?
Las células usan la transducción de señales para realizar una serie de reacciones químicas muy rápidas. Estas reacciones son llevadas a cabo por proteínas especiales llamadas enzimas o por otras sustancias que actúan como "segundos mensajeros". Todo esto ocurre en fracciones de segundo o en unos pocos segundos.
Cuando una señal llega a una célula, se produce una "cascada de señalización". Esto significa que un pequeño mensaje inicial puede generar una respuesta muy grande dentro de la célula. Es como si una pequeña chispa encendiera un gran fuego.
Pero más allá de amplificar las señales, estas vías de comunicación ayudan a las células a controlar muchas de sus funciones. Por ejemplo, deciden cuándo crecer, cuándo dividirse, cuándo madurar o incluso cuándo deben desaparecer si ya no son necesarias.
En organismos muy pequeños, como las bacterias, la transducción de señales les permite reaccionar a lo que sucede a su alrededor. En organismos más grandes, como nosotros, las células responden a muchas señales químicas. Algunas de estas señales, como las hormonas, son producidas por el propio cuerpo. Otras vienen de fuera, como el oxígeno o los nutrientes que obtenemos de los alimentos.
A pesar de la gran variedad de señales, la naturaleza ha desarrollado solo unas pocas formas principales en que las células reciben y procesan estos mensajes. Esto se ve en los receptores celulares.
Los receptores celulares tienen dos partes importantes:
- Una parte que reconoce y detecta la señal.
- Otra parte que inicia la acción dentro de la célula.
Al final de esta cadena de comunicación, las células tienen "maquinarias" especiales que generan la respuesta. Cada tipo de célula tiene sus propias maquinarias, por lo que la misma señal puede causar una respuesta diferente en distintos tipos de células. Por ejemplo, una señal para crecer puede hacer que una célula de la piel se divida, mientras que la misma señal podría hacer que una célula muscular se fortalezca.
En resumen, la forma en que los seres vivos detectan y responden a los estímulos depende de estas señales de transducción dentro de sus células.
¿Qué activa a las células?
El ambiente que rodea a una célula puede afectarla de muchas maneras:
- Diferentes tipos de moléculas pueden interactuar con su superficie.
- La temperatura puede calentarla o enfriarla.
- La radiación electromagnética (como la luz) puede activarla.
- Las células pueden estirarse, encogerse o cargarse eléctricamente, como las células musculares y las neuronas.
Las señales de transducción son clave para que las células respondan a cada uno de estos estímulos. Muchas señales vienen de fuera de la célula y se unen a la membrana plasmática. Por ejemplo, los neurotransmisores, que permiten que las neuronas se comuniquen, se unen a receptores en la membrana y abren "canales" que permiten el paso de iones.
¿Qué hacen las células en respuesta?
Las respuestas que se activan por las señales de transducción incluyen:
- Controlar la expresión genética, es decir, qué genes se "encienden" o "apagan".
- Regular el metabolismo, como la forma en que la célula produce energía.
- Permitir el movimiento de la célula, cambiando su estructura interna.
Cuando se activan los genes, se pueden producir muchas proteínas nuevas. Algunas de estas proteínas son enzimas, otras son "factores de transcripción" que pueden activar aún más genes. Así, una señal inicial puede activar un programa completo de genes y una gran variedad de cambios en la célula. Un ejemplo de esto es el inicio del desarrollo de un nuevo ser vivo después de la fecundación.
Tipos de mensajes celulares
Mensajes de fuera de la célula
En la transducción de señales, las moléculas de señalización que vienen de fuera de la célula (llamadas ligandos) se unen a receptores en la superficie de la membrana plasmática. Esta unión provoca un cambio en la forma del receptor, lo que inicia los eventos dentro de la célula. Los receptores suelen ser muy específicos: solo responden a una molécula en particular, o a una muy parecida.
Hay muchas moléculas que pueden llevar información de un lugar a otro:
- Aminoácidos y sus derivados, como la adrenalina, que actúan como neurotransmisores y hormonas.
- Gases, como el óxido nítrico.
- Los esteroides, que son hormonas que regulan el desarrollo, el metabolismo y otras funciones.
- Eicosanoides, que controlan procesos como el dolor y la inflamación.
- Una gran variedad de polipéptidos y proteínas, que regulan la división celular, la maduración y la respuesta del sistema de defensas del cuerpo.
La mayoría de las veces, estas moléculas de señalización externas son reconocidas por receptores específicos en la superficie de la célula. Estos receptores se unen fuertemente a las moléculas y transforman esa interacción externa en cambios internos.
Algunos tipos importantes de receptores son:
- Receptores acoplados a proteínas G (GPCR): Son una gran familia de receptores que atraviesan la membrana siete veces. Cuando una molécula se une a ellos, activan unas proteínas especiales llamadas proteínas G.
- Tirosina quinasas receptoras (RTK): Son otro tipo de receptores que, al unirse a una molécula, añaden grupos fosfato a otras proteínas dentro de la célula, activándolas. Son importantes para el crecimiento y la supervivencia de las células.
- Canales iónicos activados por ligando: Estos receptores son como puertas en la membrana. Cuando una molécula se une a ellos, la puerta se abre y permite el paso de iones (partículas con carga eléctrica) dentro o fuera de la célula, lo que transmite la señal. Esto es fundamental en las neuronas.
- Receptores para hormonas esteroideas: A diferencia de los anteriores, estos receptores no están en la superficie, sino dentro de la célula (en el citoplasma). Las hormonas esteroideas pueden atravesar la membrana y unirse a estos receptores. Una vez unidas, el complejo hormona-receptor se mueve al núcleo celular y controla la actividad de ciertos genes.
Mensajes dentro de la célula
Los eventos que ocurren dentro de la célula después de que una señal externa es recibida se consideran parte de la transducción de señal. Las moléculas que transmiten estos mensajes internos se llaman a menudo "segundos mensajeros".
Algunos ejemplos de segundos mensajeros son:
- AMP cíclico (AMPc) y GMP cíclico (GMPc).
- Iones de calcio.
- Moléculas derivadas de fosfolípidos, como el diacilglicerol (DAG) y el inositoltrifosfato (IP3).
- Varias proteínas quinasas y fosfatasas.
Mensajes entre células
La comunicación entre células es vital en organismos complejos. En los animales, esta comunicación se divide en varios tipos:
- Señalización endócrina: Las hormonas son producidas por células especiales y viajan por la sangre a todo el cuerpo. Es una respuesta lenta, de larga duración y actúa a distancia.
- Señalización paracrina: Las señales actúan solo en células cercanas a las que las producen, como los neurotransmisores en las sinapsis. Es una respuesta local.
- Señalización autocrina: La célula que produce la sustancia es la misma que la recibe. Un ejemplo se ve en las células del sistema de defensas del cuerpo.
- Señales yuxtacrinas: Las señales se transmiten directamente entre células que están en contacto físico, a través de proteínas en sus membranas.
Hormonas
Las hormonas son las principales moléculas que permiten la comunicación entre células o tejidos en animales y plantas. El proceso de señalización hormonal sigue varios pasos:
- La hormona se fabrica.
- Se almacena y se libera.
- Viaja hasta la célula a la que debe llegar.
- La hormona es reconocida por un receptor especial, lo que cambia la forma del receptor.
- Se inicia y amplifica la señal dentro de la célula, causando reacciones químicas. A veces, la respuesta de la célula puede enviar una señal de vuelta para que se deje de producir la hormona.
- La hormona es eliminada.
Una célula puede tener varios receptores diferentes que reconocen la misma hormona, pero que activan distintas vías de señalización. Además, diferentes tipos de tejidos pueden responder de manera distinta a la misma hormona.
Tipos de receptores celulares
Receptores en la membrana
Los receptores transmembrana son proteínas que atraviesan la membrana plasmática de la célula. Tienen una parte fuera de la célula y otra dentro. Cuando la parte externa reconoce una hormona, todo el receptor cambia de forma, lo que activa la parte interna. La hormona no entra en la célula. A menudo, la unión de la hormona hace que varias moléculas receptoras se unan entre sí.
Reconocimiento de la hormona
La hormona se une al receptor mediante enlaces débiles, como los puentes de hidrógeno. La fuerza de la señal que se transmite depende de cuántos complejos hormona-receptor se formen. Esto, a su vez, depende de la afinidad entre la hormona y el receptor, y de la cantidad de hormona y receptores disponibles.
La célula también puede ajustar la sensibilidad del receptor o cambiar el número de receptores para modificar la fuerza de la señal.
Transmisión de señales por cambios en los receptores
La transmisión de señales a través de la membrana solo es posible si muchos componentes trabajan juntos. Primero, el receptor reconoce la hormona. Luego, activa otras proteínas dentro de la célula gracias a un cambio en su forma. Estas proteínas activadas suelen estar cerca de la membrana o unidas a ella. Finalmente, envían la señal al interior de la célula.
Receptores con actividad tirosina quinasa
Este grupo incluye los receptores de la mayoría de los factores de crecimiento y de la insulina. Tienen una parte que se une a la señal, una parte que atraviesa la membrana y una parte interna con actividad de "tirosina quinasa". Cuando la señal se une, el receptor se une a otro igual (se dimeriza) y se "auto-fosforila" (se añade grupos fosfato a sí mismo). Esto activa la tirosina quinasa, que a su vez activa muchas otras moléculas en cascada. Estos receptores pueden activar diferentes vías de señalización, como las que controlan el crecimiento y la supervivencia celular.
Receptores que activan quinasas externas
Aquí se encuentran los receptores de muchas citoquinas (moléculas del sistema de defensas), como las interferones y la hormona del crecimiento. Cuando estos receptores reciben una señal, activan unas quinasas llamadas JAK. Estas quinasas activan a su vez unos factores de transcripción llamados STATs, que se mueven al núcleo y activan genes específicos.
Receptores asociados a proteínas G
Estos receptores tienen siete partes que atraviesan la membrana y están conectados a unas proteínas llamadas proteínas G. Son la familia más grande de receptores. Muchos medicamentos comunes actúan sobre ellos. Cuando una señal se une, el receptor cambia de forma y activa la proteína G, que se divide en dos partes activas. Estas partes regulan otras moléculas dentro de la célula, a veces usando el AMP cíclico o el calcio como segundos mensajeros.
Receptores que son canales iónicos
Un canal iónico activado por una señal puede reconocer otras moléculas. Después de cambiar su forma, abre un canal en la membrana por donde pueden pasar iones. Estos iones son los que transmiten la señal. Un ejemplo de esto ocurre en las células que reciben señales a través de las sinapsis.
Receptores que cambian el potencial de la membrana
Un canal iónico también puede abrirse cuando el receptor es activado por un cambio en la carga eléctrica de la célula (el potencial de membrana). Si esto ocurre, el canal iónico se abre y permite el paso de iones. En las neuronas, este mecanismo es la base del impulso nervioso que viaja a lo largo de ellas.
Receptores dentro del núcleo
Los receptores nucleares o citoplasmáticos son proteínas que se encuentran en el citoplasma o en el núcleo celular. La hormona, que suele ser una sustancia grasa, atraviesa la membrana de la célula, llega al receptor e inicia la cascada de señales. Estos receptores, una vez activados por la hormona, se mueven al núcleo y activan la transcripción de ciertos genes, lo que lleva a la producción de una proteína.
Las hormonas típicas de estos receptores son las hormonas esteroideas (como la testosterona y la progesterona) y las vitaminas A y D. Estas hormonas son muy importantes para el metabolismo, el desarrollo y la maduración de los organismos. La fuerza de la señal depende de la concentración de la hormona, que está regulada por:
- La producción y liberación de hormonas por órganos especiales (como el hipotálamo y la hipófisis).
- La disponibilidad de la hormona dentro de la célula.
- La modificación de las hormonas en la célula.
Los receptores nucleares activados por hormonas se unen a secuencias específicas de ADN llamadas "elementos sensibles a hormonas" (HREs). Como este complejo activa la transcripción de genes, estas hormonas también se llaman "inductores de la expresión genética". Los efectos de estas hormonas suelen ser a largo plazo, ya que implican la producción de nuevas proteínas.
Receptores esteroideos
Los receptores esteroideos son un tipo de receptores nucleares que se encuentran en el citoplasma. Cuando no hay hormona esteroidea, están unidos a unas proteínas llamadas "chaperonas" o "proteínas de choque térmico" (HSPs). Estas HSPs son necesarias para que el receptor cambie de forma y pueda unirse al ADN.
Amplificación de la señal
Un principio clave de la transducción de señales es la amplificación. Por ejemplo, la unión de una o pocas moléculas de neurotransmisores puede activar la entrada de millones de iones en una neurona. La unión de una o varias hormonas puede iniciar una reacción en cadena que afecta a muchas rutas metabólicas. La amplificación puede ocurrir en varios puntos de la vía de señalización.
Amplificación de la señal del receptor hormonal
Un receptor activado por una hormona puede activar muchas proteínas dentro de la célula. Por ejemplo, una molécula de rodopsina en el ojo, activada por un fotón de luz, puede activar hasta 2000 moléculas efectoras por segundo. La fuerza total de la amplificación de la señal por un receptor depende de:
- El tiempo que el complejo hormona-receptor permanece activo.
- La rapidez con la que las proteínas efectoras se activan y se separan del receptor.
- La desactivación del receptor activado.
Transducción de señal dentro de la célula
La transducción de señal dentro de la célula es realizada principalmente por moléculas llamadas segundos mensajeros.
El calcio como segundo mensajero
El calcio actúa como una molécula de señal dentro de la célula. Cuando el calcio es liberado de los almacenes internos (como el retículo endoplásmico), actúa rápidamente y en un espacio limitado. Normalmente, la concentración de iones de calcio dentro de la célula es muy baja.
El calcio está involucrado en muchos procesos, como la contracción muscular, la liberación de neurotransmisores, la visión, el crecimiento, la secreción, el movimiento celular y la actividad de los genes. El calcio puede regular proteínas de dos maneras:
- Reconociendo directamente el calcio.
- Uniéndose al centro activo de una enzima.
Una de las interacciones más estudiadas del calcio es con la calmodulina. El complejo calcio-calmodulina es importante en el crecimiento, la división celular y la señalización en las neuronas.
El óxido nítrico como segundo mensajero
El gas óxido nítrico es una molécula que puede difundirse a través de la membrana de la célula y afectar a las células vecinas. Se forma a partir de la arginina y el oxígeno. El óxido nítrico actúa principalmente activando una enzima llamada guanilato ciclasa, que produce otro segundo mensajero, el guanosín monofosfato cíclico (GMPc).
El óxido nítrico tiene tres funciones principales:
- Relajar los vasos sanguíneos.
- Regular la liberación de neurotransmisores.
- Participar en la respuesta de las defensas del cuerpo.
Galería de imágenes
-
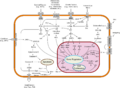
Vista general de los patrones de transducción de señales.

Véase también
 En inglés: Signal transduction Facts for Kids
En inglés: Signal transduction Facts for Kids
