
Clasificación de los virus para niños
La clasificación de los virus es como organizar una gran biblioteca de seres diminutos. Se trata de darles nombres y agruparlos en un sistema, parecido a cómo clasificamos a los animales o las plantas.
Los virus se clasifican principalmente por sus características visibles y funcionales. Por ejemplo, se mira la forma de su cápside (su cubierta protectora), el tipo de ácido nucleico que tienen (si es ADN o ARN), las proteínas que los forman, cómo se multiplican dentro de una célula, a qué organismos infectan y qué tipo de enfermedad pueden causar.
Existen dos sistemas principales para clasificar los virus. Uno es el del Comité Internacional de Taxonomía de Virus (ICTV), que es el sistema oficial y establece las reglas para nombrar y clasificar los virus. El otro es el sistema de clasificación de Baltimore, que los agrupa en siete categorías según cómo producen su ARNm (una molécula esencial para crear proteínas). Estos dos sistemas no compiten, ¡al contrario! Se complementan y el ICTV incluso usa algunos criterios del sistema de Baltimore.
A diferencia de los seres vivos, que se cree que vienen de un ancestro común, los virus tienen muchos orígenes diferentes. Por eso, su clasificación no sigue una única línea evolutiva.
Contenido
- ¿Por qué los virus no están en el árbol de la vida?
- ¿Cómo se clasifican los virus por su forma?
- ¿Qué es la clasificación de Baltimore?
- ¿Qué organismos pueden infectar los virus?
- Agentes subvíricos
- Clasificación del ICTV
- Clasificación del ICTV (2025)
- Galería de imágenes
- Véase también
¿Por qué los virus no están en el árbol de la vida?
Según la teoría celular, los virus no se consideran seres vivos. La mayoría de los científicos los excluyen de los sistemas de clasificación biológica y del "árbol de la vida", que muestra cómo se relacionan evolutivamente los seres vivos. Esto se debe a que los virus son muy simples y cambian rápidamente.
El ICTV y muchos científicos ven a los virus como "elementos genéticos móviles". Esto significa que son como pequeños paquetes de información genética que se mueven. No cumplen con todas las funciones que definen la vida, como tener su propia energía o metabolismo, o poder reproducirse y evolucionar por sí solos. Solo pueden hacerlo dentro de las células de otros organismos. Sin una célula huésped, los virus son como "materia orgánica compleja sin vida".
Aquí te explicamos algunas razones por las que los virus no se consideran vivos y no se incluyen en el árbol de la vida:
- Los virus no tienen energía ni metabolismo propio. No pueden reproducirse ni evolucionar sin una célula huésped.
- Los virus no comparten características con las células, y no hay un solo gen que todos los virus tengan en común.
- No se ha encontrado un gen ancestral que sea compartido por todos los virus.
- Es difícil saber cuándo aparecieron los primeros virus, porque pueden pasar fácilmente de un huésped a otro.
- Los virus no heredan estructuras como las membranas de las células; todo lo que tienen lo fabrican de nuevo.
- Los genes que los virus usan para el metabolismo vienen de las células que infectan, no al revés.
- Los virus "toman prestados" genes de las células, no los inventan ni los dan a las células. La mayoría de los genes se mueven de las células a los virus.
- Aunque muchos virus son simples, esto no significa que sean primitivos. Su tamaño pequeño les ayuda a multiplicarse rápido.
A pesar de no ser considerados vivos, los virus son muy importantes. Han influido mucho en la evolución de la vida en la Tierra, ya que obligan a los organismos a desarrollar defensas. También pueden transferir genes entre organismos, lo que aumenta la variedad genética.
¿Cómo se clasifican los virus por su forma?
Los virus también se pueden clasificar por su forma, que puede ser:
- Virus icosaedro: Tienen una forma parecida a un dado de 20 caras. Ejemplos: adenovirus, herpesvirus.
- Virus filamentoso o helicoidal: Son largos y delgados, como un hilo o un muelle. Ejemplos: virus del mosaico del tabaco, virus del Ébola.
- Virus ovoide: Tienen forma de huevo. Ejemplos: algunos virus gigantes.
- Virus inusual: Tienen formas muy extrañas, como de limón, lágrima o espiral. Ejemplos: rhabdovirus, muchos virus de arqueas.
- Virus sin cápside: Son virus de ARN que no tienen la cubierta protectora (cápside). Se encuentran principalmente en hongos y protistas.
¿Qué es la clasificación de Baltimore?

David Baltimore, un biólogo que ganó el Premio Nobel, creó este sistema de clasificación. Se usa junto con el sistema del ICTV para clasificar los virus hoy en día.
La clasificación de Baltimore se basa en cómo los virus producen su ARNm. Los virus necesitan crear ARNm a partir de su material genético para fabricar proteínas y multiplicarse. Pero cada tipo de virus lo hace de una manera diferente. El material genético de los virus puede ser de una sola cadena (ss) o de doble cadena (ds), y puede ser de ARN o ADN. Algunos usan una enzima especial llamada transcriptasa inversa. Además, los virus de ARN de una sola cadena pueden ser de sentido positivo (+) o negativo (–).
Esta clasificación divide los virus en siete grupos:
- I: Virus ADN bicatenario (ADN de doble cadena)
- II: Virus ADN monocatenario (ADN de una sola cadena)
- III: Virus ARN bicatenario (ARN de doble cadena)
- IV: Virus ARN monocatenario positivo (ARN de una sola cadena, sentido positivo)
- V: Virus ARN monocatenario negativo (ARN de una sola cadena, sentido negativo)
- VI: Virus ARN monocatenario retrotranscrito (ARN de una sola cadena que usa transcriptasa inversa)
- VII: Virus ADN bicatenario retrotranscrito (ADN de doble cadena que usa transcriptasa inversa)
El tipo de material genético y cómo se multiplican los virus varía mucho entre estos grupos.
Virus de ADN
La mayoría de los virus ADN se multiplican en el núcleo de la célula que infectan. Si la célula tiene la "puerta" adecuada en su superficie, el virus entra. Estos virus dependen mucho de la maquinaria de la célula para copiar su ADN y ARN. El material genético del virus debe llegar al núcleo para usar esta maquinaria.
Virus de ADN de doble cadena
Los virus de ADN de doble cadena tienen su material genético formado por ADN de doble cadena. Para multiplicarse, necesitan entrar en el núcleo de la célula huésped y usar las herramientas de la célula. Estos virus dependen de que la célula esté en una fase de multiplicación activa. Pueden incluso forzar a la célula a dividirse, lo que a veces puede llevar a cambios en la célula. Infectan a animales, protistas, hongos, bacterias y arqueas.
Virus de ADN de una sola cadena
Los virus de ADN de una sola cadena tienen ADN de una sola cadena. Para multiplicarse, su ADN de una sola cadena debe convertirse en ADN de doble cadena dentro de la célula infectada. Infectan a todo tipo de organismos celulares, como animales, plantas, hongos, protistas, bacterias y arqueas.
Virus de ARN
Los virus ARN son especiales porque su información genética está en ARN. Esto significa que usan el ARN como su material genético o lo necesitan para multiplicarse. Su multiplicación suele ocurrir en el citoplasma de la célula. Los virus de ARN usan su propia enzima, llamada ARN polimerasa dependiente de ARN, para hacer copias de su material genético.
Virus de ARN de doble cadena
Los virus de ARN de doble cadena tienen ARN de doble cadena en su material genético. Se multiplican en el citoplasma y no dependen de las herramientas de la célula huésped como los virus de ADN, porque traen sus propias enzimas. Infectan a animales, plantas, hongos, protistas y bacterias.
Virus de ARN de una sola cadena, sentido positivo
Los virus de ARN de una sola cadena, sentido positivo tienen ARN que es como un "mensaje" listo para ser leído por la célula huésped. Pueden empezar a producir proteínas de inmediato. Su multiplicación ocurre principalmente en el citoplasma. Infectan a todos los organismos celulares, siendo más comunes en animales y plantas.
Virus de ARN de una sola cadena, sentido negativo
Los virus de ARN de una sola cadena, sentido negativo tienen ARN que es el "complemento" del mensaje. Primero, necesitan una enzima especial (que el virus trae consigo) para convertir su ARN en ARN de sentido positivo, que luego la célula puede leer. Infectan a todos los organismos celulares, siendo más comunes en animales y plantas.
Virus retrotranscritos
Los virus retrotranscritos son un tipo especial que usan un proceso llamado "transcripción inversa". Esto significa que pueden crear ADN a partir de una plantilla de ARN.
Virus de ARN de una sola cadena retrotranscrito
Un virus ARN monocatenario retrotranscrito usa una enzima llamada transcriptasa inversa para crear ADN a partir de su genoma de ARN. Este ADN a menudo se integra en el material genético de la célula huésped, como en el caso de los retrovirus. Infectan a todo tipo de eucariotas (organismos con células complejas), como animales, plantas, hongos y protistas.
Virus de ADN de doble cadena retrotranscrito
Estos virus se multiplican usando la transcripción inversa, creando ADN a partir de una plantilla de ARN. Este grupo incluye virus que infectan a animales (como los de la familia Hepadnaviridae) y a plantas (como los de la familia Caulimoviridae).
¿Qué organismos pueden infectar los virus?
Los virus pueden infectar a todo tipo de organismos. Aunque están en todo el mundo, cada tipo de virus suele infectar a un grupo específico de organismos.
Virus que infectan células
Virus de bacterias
Los bacteriófagos son virus que infectan bacterias. Son muy comunes, especialmente en el agua. Se unen a la superficie de las bacterias y entran en ellas. En poco tiempo, las herramientas de la bacteria empiezan a producir nuevas partículas de virus. Las bacterias se defienden produciendo enzimas que destruyen el ADN extraño del virus. También tienen un sistema llamado CRISPR que les da una especie de "memoria" para defenderse de virus que ya han encontrado antes.
Virus de arqueas
Así como los bacteriófagos infectan bacterias, algunos virus se multiplican dentro de las arqueas (otro tipo de microorganismos). Estos virus pueden tener formas conocidas o muy inusuales, como botellas o lágrimas. Las arqueas también se defienden con sistemas parecidos a los de las bacterias.
Virus de protistas
Estos virus pueden infectar a diversos organismos eucariotas simples como algas, protozoos y mohos mucilaginosos. Se sabe poco de ellos, pero pueden ser importantes para controlar las poblaciones de algas y protozoos. Algunos de los virus más grandes conocidos infectan principalmente amebas.
Virus de hongos
Los virus que infectan hongos (micovirus) pueden tener efectos variados, desde no causar síntomas hasta ser perjudiciales. La mayoría no causan problemas. Se cree que se mueven dentro del hongo a través del flujo de su plasma.
Virus de plantas
Hay muchos tipos de virus de las plantas o fitovirus. A menudo, solo causan una reducción en la producción de cultivos. Suelen ser transmitidos por insectos, pero también por algunos hongos o nemátodos. Las plantas tienen defensas elaboradas contra los virus, como los "genes de resistencia" que causan la muerte de las células infectadas para detener la expansión del virus. También producen sustancias naturales que combaten los virus.
Virus de animales
Los virus son importantes causantes de enfermedades en el ganado. Por ejemplo, la fiebre aftosa es causada por un virus. Las mascotas, como perros y gatos, también pueden sufrir infecciones virales graves si no están vacunadas. Afortunadamente, la mayoría de los virus coexisten sin causar daño a sus huéspedes.
Virus con más de dos tipos de huéspedes diferentes
Algunos virus pueden infectar a dos o más tipos de huéspedes diferentes. Por ejemplo, algunos virus pueden infectar animales, plantas, hongos y protistas, incluyendo a los ser humanos. Los caudovirus, que son bacteriófagos, no solo infectan bacterias, sino también arqueas.
Virus satélite
Aunque los virus dependen de las células para vivir, existen otros virus que dependen de otros virus para multiplicarse. Estos se llaman virus satélite. Son más pequeños que los virus comunes y necesitan la ayuda de un "virus auxiliar" para poder replicarse. Generalmente, no dañan a sus virus auxiliares. Sin embargo, algunos, como los virófagos, sí pueden ser perjudiciales para sus virus auxiliares, impidiendo que se multipliquen o causando malformaciones. Los virus satélite también pueden ayudar a las células huésped a defenderse de los virus que las infectan.
Agentes subvíricos


Los agentes subvíricos son partículas biológicas que se parecen a los virus, pero no cumplen con todas sus características típicas. Se consideran más simples que los virus.
Un virus típico tiene material genético (ADN o ARN) cubierto por una cápside. Un agente subvírico no cumple completamente con esta definición. Algunos ejemplos son:
- Satélite: Un virus que necesita de otro virus para multiplicarse dentro de una célula.
- Viroide: Un virus que no tiene cápside; su material genético (ARN) está "desnudo".
- Obelisco: Un tipo de elemento genético móvil con una forma particular.
- Virusoide: Similar a un viroide (ARN desnudo), pero también necesita de otro virus para multiplicarse.
- Elemento vírico endógeno: Una secuencia genética de virus que se ha integrado en el material genético de una célula.
- Partícula defectiva interferente: Una parte de ARN o ADN de virus que se forma por un error y reduce la capacidad de infección del virus.
- Partícula similar a virus: Una parte de las proteínas de un virus, sin material genético, que se usa en vacunas.
- Prion: Un agente infeccioso formado solo por proteínas que pueden cambiar de forma y causar enfermedades.
El ICTV reconoce a los viroides, satélites y virusoides en su clasificación. Los priones también son reconocidos, pero se clasifican de forma separada.
Clasificación del ICTV
El Comité Internacional de Taxonomía de Virus (ICTV) empezó a crear reglas para nombrar y clasificar los virus en los años 70. Es el único organismo encargado de mantener una clasificación universal de virus. Su sistema es similar al de los organismos celulares, pero tiene algunas diferencias.
La clasificación de los virus comienza en el nivel de Dominio y sigue así, con los sufijos que los identifican:
- Dominio (-viria)
- Reino (-virae)
- Filo (-viricota)
- Subfilo (-viricotina)
- Clase (-viricetes)
- Orden (-virales)
- Suborden (-virineae)
- Familia (-viridae)
- Subfamilia (-virinae)
- Género (-virus)
- Especie (-virus)
A diferencia de los seres vivos, que tienen un nombre científico de dos partes (como Homo sapiens), los nombres de las especies de virus no tienen una forma estándar. El ICTV pide que el nombre de una especie sea lo más corto y claro posible. A menudo, los nombres de las especies de virus se forman con el nombre de la enfermedad que causan.
¿Qué es una especie de virus?
Una especie de virus es un grupo de virus que tienen propiedades únicas y que se pueden distinguir de otras especies. Son como una "familia" de virus que se multiplican y viven en un lugar específico.
Los virus son entidades físicas que evolucionan, mientras que las especies y las categorías superiores son conceptos que usamos para entenderlos.
Definición del ICTV para cada entidad vírica
El ICTV define los virus y otras entidades relacionadas de la siguiente manera:
- Virus: Son un tipo de elemento genético móvil que codifica al menos una proteína principal de su cubierta (virión).
- Viroides: Son elementos genéticos móviles de ARN de una sola cadena, pequeños y sin cubierta. No codifican proteínas y no necesitan virus para transmitirse. Se multiplican solos.
- Satélites/Virusoides: Son elementos genéticos móviles que no son viroides, pero que dependen de los virus para multiplicarse y transmitirse.
Los virus, viroides y virusoides/satélites tienen una historia evolutiva relacionada con otros elementos genéticos móviles.
Dominios de virus
La clasificación actual del ICTV se basa en el tipo de proteínas y la secuencia de aminoácidos que tienen los virus. Los virus se dividen en 6 dominios y algunos reinos. Las ideas más aceptadas sobre el origen de los virus combinan dos hipótesis: la del "escape" (que los virus escaparon de las células) y la del "virus primero" (que los virus existieron antes que las células).
A continuación, se describen los dominios de virus según sus características y evolución:
Dominios de ARN

- Riboviria: Incluye virus que tienen enzimas especiales para copiar ARN (ARN polimerasas dependientes de ARN) o para hacer ADN a partir de ARN (transcriptasas inversas). Se cree que estas enzimas tienen un origen común. Este dominio incluye la mayoría de los virus de ARN y los virus retrotranscritos.
- Orthornavirae: Incluye la mayoría de los virus de ARN. Tienen una enzima que les permite copiar ARN sin necesidad de ADN. Se cree que el ancestro de estos virus fue una molécula de ARN simple sin cápside.
- Pararnavirae: Incluye los virus retrotranscritos. Tienen una enzima llamada transcriptasa inversa. Se cree que se originaron cuando un elemento genético móvil se integró en la cápside de un virus de ARN.
- Ribozyviria: Incluye a los virusoides, que son como virus satélites que dependen de virus de ARN o retrotranscritos para multiplicarse. Se caracterizan por ser moléculas de ARN que se copian a sí mismas usando otras moléculas de ARN llamadas ribozimas.
- Viroidia: Incluye a los viroides, que son agentes infecciosos de ARN sin cápside ni enzimas propias. No codifican proteínas y no usan ADN. Infectan plantas y recientemente se han encontrado en hongos y bacterias. Se consideran "reliquias" de las primeras formas de vida basadas en ARN.
Dominios de ADN

- Duplodnaviria: Incluye virus de ADN con una proteína específica llamada HK97-MCP y una enzima única llamada terminasa. Son virus de ADN de doble cadena con cápsides de 20 caras. Se cree que se originaron antes del último ancestro común universal (LUCA).

- Adnaviria: Incluye virus de arqueas con una proteína específica llamada SIRV2, forma filamentosa y ADN en forma de A. Son virus de ADN de doble cadena. También se cree que se originaron antes de LUCA.

- Varidnaviria: Incluye virus de ADN con una proteína específica llamada DJR-MCP. Son virus de ADN de doble cadena con cápsides de 20 caras. Se cree que se originaron antes de LUCA.
- Bamfordvirae: Incluye virus de bacterias y eucariotas. Los virus gigantes evolucionaron de virus más pequeños, aumentando su tamaño y genoma.
- Abadenavirae: Similares a los anteriores, pero más primitivos y solo infectan procariotas.

- Singelaviria: Incluye virus de ADN de doble cadena con cápsides de 20 caras y una proteína específica llamada SJR-MCP. Solo infectan procariotas. Se cree que se originaron antes de LUCA.
- Monodnaviria: Incluye virus de ADN con una proteína específica llamada rep que realiza una replicación especial. Casi todos son virus de ADN de una sola cadena. Se cree que se originaron a partir de elementos genéticos móviles de bacterias que se integraron en virus de ARN.
- Shotokuvirae: Contiene virus de eucariotas como los papilomavirus y parvovirus.
- Sangervirae: Contiene virus de bacterias.
- Trapavirae: Contiene virus de arqueas.
- Loebvirae: Contiene virus de procariotas con cápsides helicoidales.
- Telodnaviria: Incluye virus de ADN de doble cadena con cápsides ovoides o filamentosas. Su origen es desconocido.
- Linaje de virus fusiformes y Clavaviridae: Incluye virus de arqueas con formas de limón o filamentosas. Se cree que están relacionados y que se originaron antes de LUCA.
Además, hay 16 familias de virus de ADN de arqueas que no se han asignado a ningún dominio porque no tienen proteínas o genes similares a otros virus. Se cree que podrían ser descendientes de virus muy antiguos que se extinguieron.
Clasificación del ICTV (2025)
Aquí te mostramos la clasificación más reciente del ICTV:
- Dominio Riboviria
- Reino Pararnavirae
- Familia Hepadnaviridae
- Orden Ortervirales
- Familia Belpaoviridae
- Familia Caulimoviridae
- Familia Retroviridae
- Familia Metaviridae
- Familia Pseudoviridae
- Reino Orthornavirae
- Filo Lenarviricota
- Familia Botourmiaviridae
- Familia Mitoviridae
- Clase Amabiliviricetes
- Filo Lenarviricota
- Reino Pararnavirae
* Familia Narnaviridae * Familia Splipalmiviridae
-
-
-
- Clase Leviviricetes
-
-
* Orden Norzivirales ** Familia Atkinsviridae ** Familia Duinviridae ** Familia Fiersviridae ** Familia Solspiviridae * Orden Timlovirales ** Familia Blumeviridae ** Familia Steitzviridae * Género Grandbuvirus * Género Mahrahovirus * Género Nicedsevirus * Género Nordovirus * Género Skrubnovirus
-
-
- Filo Duplornaviricota
- Familia Cystoviridae
- Orden Reovirales
- Filo Duplornaviricota
-
* Familia Sedoreoviridae * Familia Spinareoviridae
-
-
-
- Orden Ghabrivirales
-
-
* Familia Alternaviridae * Suborden Alphatotivirineae ** Familia Botybirnaviridae ** Familia Chrysoviridae ** Familia Fusagraviridae ** Familia Megabirnaviridae ** Familia Monocitiviridae ** Familia Orthototiviridae ** Familia Phlegiviridae ** Familia Pseudototiviridae ** Familia Quadriviridae ** Familia Spiciviridae * Suborden Betatotivirineae ** Familia Artiviridae ** Familia Giardiaviridae ** Familia Inseviridae ** Familia Lebotiviridae ** Familia Megatotiviridae ** Familia Ootiviridae ** Familia Pistolviridae ** Familia Yadonushiviridae
-
-
- Filo Negarnaviricota
- Subfilo Haploviricotina
- Filo Negarnaviricota
-
* Familia Qinviridae * Familia Aspiviridae * Familia Yueviridae * Clase Monjiviricetes ** Orden Jingchuvirales *** Familia Chuviridae *** Familia Aliusviridae *** Familia Chuviridae *** Familia Crepuscuviridae *** Familia Myriaviridae *** Familia Nataviridae ** Orden Mononegavirales *** Familia Artoviridae *** Familia Bornaviridae *** Familia Filoviridae *** Familia Lispiviridae *** Familia Mymonaviridae *** Familia Nyamiviridae *** Familia Paramyxoviridae *** Familia Pneumoviridae *** Familia Rhabdoviridae *** Familia Sunviridae *** Familia Xinmoviridae
-
-
-
- Subfilo Polyploviricotina
-
-
* Clase Bunyaviricetes ** Orden Elliovirales *** Familia Cruliviridae *** Familia Fimoviridae *** Familia Hantaviridae *** Familia Peribunyaviridae *** Familia Phasmaviridae *** Familia Tospoviridae *** Familia Tulasviridae ** Orden Hareavirales *** Familia Arenaviridae *** Familia Discoviridae *** Familia Konkoviridae *** Familia Leishbuviridae *** Familia Mypoviridae *** Familia Nairoviridae *** Familia Phenuiviridae *** Familia Tosoviridae *** Familia Wupedeviridae
-
-
-
- Clase Insthoviricetes
-
-
* Orden Articulavirales ** Familia Amnoonviridae ** Familia Orthomyxoviridae
-
-
- Filo Kitrinoviricota
- Familia Flaviviridae
- Orden Nodamuvirales
- Filo Kitrinoviricota
-
* Familia Nodaviridae * Familia Sinhaliviridae
-
-
-
- Orden Tolivirales
-
-
* Familia Carmotetraviridae * Familia Tombusviridae
-
-
-
- Clase Alsuviricetes
-
-
* Orden Hepelivirales ** Familia Alphatetraviridae ** Familia Benyviridae ** Familia Hepeviridae ** Familia Matonaviridae * Orden Martellivirales ** Familia Bromoviridae ** Familia Closteroviridae ** Familia Endornaviridae ** Familia Kitaviridae ** Familia Mayoraviridae ** Familia Togaviridae ** Familia Virgaviridae * Orden Tymovirales ** Familia Alphaflexiviridae ** Familia Betaflexiviridae ** Familia Deltaflexiviridae ** Familia Gammaflexiviridae ** Familia Tymoviridae
-
-
- Filo Ambiviricota
- Familia Dumbiviridae
- Familia Quambiviridae
- Familia Trimbiviridae
- Familia Unambiviridae
- Filo Pisuviricota
- Clase Stelpaviricetes
- Filo Ambiviricota
-
* Familia Astroviridae * Familia Potyviridae
-
-
-
- Clase Duplopiviricetes
-
-
* Orden Durnavirales ** Familia Amalgaviridae ** Familia Curvulaviridae ** Familia Hypoviridae ** Familia Fusariviridae ** Familia Partitiviridae ** Familia Picobirnaviridae ** Familia Soropartitiviridae
-
-
-
- Clase Pisoniviricetes
-
-
* Orden Nidovirales ** Familia Coronaviridae ** Familia Mononiviridae ** Familia Tobaniviridae ** Suborden Adnidovirinae *** Familia Abyssoviridae *** Familia Arteriviridae *** Familia Cremegaviridae *** Familia Gresnaviridae *** Familia Olfoviridae ** Suborden Nanivirinae *** Familia Nanghosviridae *** Familia Nanhypoviridae ** Suborden Mesnidovirinae *** Familia Medioniviridae *** Familia Mesoniviridae ** Suborden Runidovirinae *** Familia Euroniviridae *** Familia Roniviridae * Orden Picornavirales ** Familia Caliciviridae ** Familia Dicistroviridae ** Familia Iflaviridae ** Familia Marnaviridae ** Familia Picornaviridae ** Familia Polycipiviridae ** Familia Noraviridae ** Familia Secoviridae ** Familia Solinviviridae * Orden Sobelivirales ** Familia Alvernaviridae ** Familia Barnaviridae ** Familia Solemoviridae * Orden Yadokarivirales ** Familia Yadokariviridae ** Familia Hadakariviridae ** Familia Polymycoviridae
-
-
-
- Clase Orpoviricetes
-
-
* Orden Bormycovirales ** Familia Alphaormycoviridae ** Familia Deltanormycoviridae * Orden Formycovirales ** Familia Betaormycoviridae ** Familia Gammaormycoviridae
-
-
-
- Familia Birnaviridae
- Familia Permutotetraviridae
-
- Familia Tonesaviridae
- Orden Tombendovirales
- Familia Pamosaviridae
- Familia Sarthroviridae
- Familia Tomosaviridae
-
- Dominio Adnaviria
- Familia Tristromaviridae
- Familia Ahmunviridae
- Orden Ligamenvirales
- Familia Lipothrixviridae
- Familia Rudiviridae
- Familia Chiyouviridae
- Familia Ungulaviridae
- Dominio Duplodnaviria
- Orden Herpesvirales
- Familia Alloherpesviridae
- Familia Herpesviridae
- Familia Malacoherpesviridae
- Clase Caudoviricetes
- Orden Autographivirales
- Familia Autonotataviridae
- Familia Autoscriptoviridae
- Familia Autosignataviridae
- Familia Autotranscriptaviridae
- Orden Crassvirales
- Familia Crevaviridae
- Familia Intestiviridae
- Familia Steigviridae
- Familia Suoliviridae
- Orden Grandevirales
- Familia Epsomviridae
- Familia Lakviridae
- Orden Pantevenvirales
- Familia Ackermannviridae
- Familia Straboviridae
- Familia Kyanoviridae
- Orden Kirjokansivirales
- Familia Flexireviridae
- Familia Haloferuviridae
- Familia Pyrstoviridae
- Familia Shortaselviridae
- Orden Juravirales
- Familia Yangangviridae
- Familia Yanlukaviridae
- Orden Methanobavirales
- Familia Anaeroviridae
- Familia Leisingerviridae
- Orden Nakonvirales
- Familia Ahpuchviridae
- Familia Ekchuahviridae
- Orden Thumleimavirales
- Familia Hafunaviridae
- Familia Halomagnusviridae
- Familia Queuoviridae
- Familia Soleiviridae
- Familia Aggregaviridae
- Familia Aliceevansviridae
- Familia Alisviridae
- Familia Andersonviridae
- Familia Arenbergviridae
- Familia Assiduviridae
- Familia Berryhillviridae
- Familia Casidaviridae
- Familia Casjensviridae
- Familia Chaseviridae
- Familia Chimalliviridae
- Familia Clermontviridae
- Familia Colingsworthviridae
- Familia Connertonviridae
- Familia Demerecviridae
- Familia Drexlerviridae
- Familia Duneviridae
- Familia Ehrlichviridae
- Familia Felixviridae
- Familia Fervensviridae
- Familia Forsetiviridae
- Familia Fredfastierviridae
- Familia Fuxiviridae
- Familia Grimontviridae
- Familia Guelinviridae
- Familia Helgolandviridae
- Familia Herelleviridae
- Familia Hodgkinviridae
- Familia Jeanschmidtviridae
- Familia Kleczkowskaviridae
- Familia Konodaiviridae
- Familia Kruegerviridae
- Familia Kunpengviridae
- Familia Lindbergviridae
- Familia Ludisviridae
- Familia Lutetiaviridae
- Familia Madisaviridae
- Familia Madridviridae
- Familia Mazoviaviridae
- Familia Mesyanzhinovviridae
- Familia Mktvariviridae
- Familia Molycolviridae
- Familia Naomviridae
- Familia Nixviridae
- Familia Orlajensenviridae
- Familia Pachyviridae
- Familia Peduoviridae
- Familia Pervagoviridae
- Familia Pootjesviridae
- Familia Pungoviridae
- Familia Rountreeviridae
- Familia Saffermanviridae
- Familia Salasmaviridae
- Familia Satyaviridae
- Familia Saparoviridae
- Familia Sarkviridae
- Familia Schitoviridae
- Familia Speroviridae
- Familia Stackebrandtviridae
- Familia Stanwillamsviridae
- Familia Suolaviridae
- Familia Toyamaviridae
- Familia Trautnerviridae
- Familia Umezonoviridae
- Familia Vandenendeviridae
- Familia Verandiviridae
- Familia Vertoviridae
- Familia Vilmaviridae
- Familia Winoviridae
- Familia Zierdtviridae
- Familia Zobellviridae
- Agentes de transferencia de genes (elementos genéticos procariotas derivados):
- Familia Bartogtaviriformidae
- Familia Brachygtaviriformidae
- Familia Rhodogtaviriformidae
- Orden Autographivirales
- Orden Herpesvirales
- Dominio Monodnaviria
- Reino Loebvirae
- Familia Inoviridae
- Familia Plectroviridae
- Familia Paulinoviridae
- Reino Sangervirae
- Familia Microviridae
- Reino Trapavirae
- Familia Pleolipoviridae
- Familia Thalassapleoviridae
- Reino Shotokuvirae
- Filo Cossaviricota
- Familia Bidnaviridae
- Familia Parvoviridae
- Clase Papovaviricetes
- Filo Cossaviricota
- Reino Loebvirae
* Familia Papillomaviridae * Familia Polyomaviridae
-
-
- Filo Cressdnaviricota
- Clase Repensiviricetes
- Filo Cressdnaviricota
-
* Familia Geminiviridae * Familia Genomoviridae * Familia Geplanaviridae
-
-
-
- Clase Arfiviricetes
-
-
* Familia Bacilladnaviridae * Familia Redondoviridae * Familia Smacoviridae * Familia Naryaviridae * Familia Draupnirviridae * Familia Oomyviridae * Familia Pecoviridae * Orden Gerdzevirales ** Familia Gandrviridae ** Familia Ouroboviridae * Orden Saturnivirales ** Familia Kanorauviridae ** Familia Mahapunaviridae * Orden Rohanvirales ** Familia Nenyaviridae ** Familia Adamviridae ** Familia Kirkoviridae * Orden Cirlivirales ** Familia Circoviridae ** Familia Vilyaviridae ** Familia Endolinaviridae * Orden Mulpavirales ** Familia Anicreviridae ** Familia Amesuviridae ** Familia Metaxyviridae ** Familia Nanoviridae ** Familia Alphasatellitidae
-
-
- Filo Commensaviricota
- Familia Anelloviridae
- Filo Commensaviricota
-
- Dominio Varidnaviria
- Reino Abadenavirae
- Familia Finnlakeviridae
- Clase Belvinaviricetes
- Familia Turriviridae
- Familia Chacviridae
- Familia Skuldviridae
- Orden Vinavirales
- Reino Abadenavirae
* Familia Corticoviridae * Familia Autolykiviridae * Familia Asemoviridae * Familia Mestraviridae * Familia Parnassusviridae
-
- Reino Bamfordvirae
- Filo Preplasmiviricota
- Familia Tectiviridae
- Subfilo Polisuviricotina
- Filo Preplasmiviricota
- Reino Bamfordvirae
* Familia Adenoviridae * Familia Eupolintoviridae * Familia Phypoliviridae * Clase Virophaviricetes ** Familia Ruviroviridae ** Familia Maviroviridae ** Familia Sputniviroviridae ** Orden Piklausovirales *** Familia Burtonviroviridae *** Familia Dishuiviroviridae *** Familia Gulliviroviridae *** Familia Omnilimnoviroviridae
-
-
- Filo Nucleocytoviricota
- Familia Yaraviridae
- Clase Pokkesviricetes
- Filo Nucleocytoviricota
-
* Familia Asfarviridae * Familia Poxviridae'
-
-
-
- Clase Megaviricetes
-
-
* Orden Algavirales ** Familia Phycodnaviridae ** Familia Mamonaviridae * Orden Imitervirales ** Familia Mimiviridae ** Familia Allomimiviridae ** Familia Mesomimiviridae ** Familia Schizomimiviridae * Orden Pimascovirales ** Familia Ascoviridae ** Familia Iridoviridae ** Familia Marseilleviridae ** Suborden Ocovirineae *** Familia Pithoviridae *** Familia Hdriviridae *** Familia Orpheoviridae
- Dominio Ribozyviria
- Familia Kolmioviridae
- Dominio Telodnaviria
- Clase Naldaviricetes
- Familia Nimaviridae
- Orden Lefavirales
- Familia Baculoviridae
- Familia Hytrosaviridae
- Familia Filamentoviridae
- Familia Nudiviridae
- Familia Polydnaviriformidae
- Clase Naldaviricetes
- Dominio Viroidia
- Familia Avsunviroidae
- Familia Pospiviroidae
- Familias de colocación incierta
- Dominio propuesto
- Familia Clavaviridae
- Virus fusiformes
- Familia Fuselloviridae
- Familia Halspiviridae
- Familia Thaspiviridae
- Familia Bicaudaviridae
- Familia Itzamnaviridae
- Familia Huangdiviridae
- Familia Eurekaviridae
- Familia Nipumfusiviridae
- Familia Ampullaviridae
- Familia Globuloviridae
- Familia Guttaviridae
- Familia Plasmaviridae
- Familia Spiraviridae
- Familia Ovaliviridae
- Familia Obscuriviridae
- Familia Tolecusatellitidae
- Dominio propuesto
- Género de colocación incierta
- Rhizidiovirus
Galería de imágenes
-
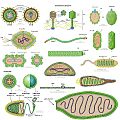
Diversos tipos de virus.

Véase también
 En inglés: Virus classification Facts for Kids
En inglés: Virus classification Facts for Kids
