
Morfogénesis para niños
La morfogénesis es un proceso biológico fascinante que explica cómo un ser vivo desarrolla su forma, desde una pequeña célula hasta un organismo completo. Es como el "origen de la forma" de todo lo que tiene vida. Este estudio es muy importante en la Biología del desarrollo, junto con el Crecimiento celular y la Diferenciación celular (cuando las células se especializan para diferentes trabajos).
La morfogénesis controla cómo las células se organizan en el espacio durante el desarrollo de un embrión. Pero no solo ocurre en los embriones; también sucede en organismos ya desarrollados, en cultivos de células o en crecimientos anormales de células. Incluso describe cómo se forman los seres vivos unicelulares o cómo ha evolucionado la forma del cuerpo de un grupo de animales a lo largo del tiempo. Esta última parte la estudia la Biología evolutiva del desarrollo.
La forma de un organismo puede cambiar por diferentes razones, como la acción de sustancias químicas especiales llamadas morfógenos, por elementos del ambiente o por cambios en el espacio donde se desarrollan las células.
Contenido
Historia de la Morfogénesis
Primeros Estudios y Patrones Naturales
Los primeros científicos que estudiaron la morfogénesis, como D'Arcy Wentworth Thompson en 1927 con su libro On growth and form, querían describir con matemáticas cómo se forman los patrones en la naturaleza. Por ejemplo, las espirales de las hojas en una planta (filotaxis) o las formas de las conchas de los caracoles.

El Papel del Núcleo Celular
En 1930, Joachim Hämmerling descubrió que el núcleo de una célula es esencial para la morfogénesis. Él usó un tipo de alga llamada Acetabularia, que tiene una forma muy particular y un solo núcleo en su base. Hämmerling trabajó con dos especies diferentes, A. mediterranea y A. crenulata.
El experimento de Hämmerling fue muy ingenioso: intercambió los núcleos de las dos especies. Observó que el "sombrerillo" (la parte superior del alga) que se formaba era el de la especie del núcleo que había sido trasplantado. Esto demostró que el núcleo de la célula controla cómo se desarrolla la forma del organismo.
La Visión de Alan Turing
Alan Turing, un famoso matemático, predijo en su libro The Chemical Basis of Morphogenesis (1952) que la morfogénesis podría explicarse por la interacción de dos señales químicas: una que estimula el crecimiento y otra que lo frena. Aunque Turing no vio estos patrones en la vida real, su idea fue muy cercana a lo que los biólogos descubrirían años después.
Los estudios de estos investigadores impulsaron otros descubrimientos importantes, como la estructura del ADN en 1953, gracias a los trabajos de Rosalind Franklin, James D. Watson y Francis Crick. Esto dio origen a la Biología molecular y la Bioquímica.
Descubrimiento de Morfógenos
En 1960, Lewis Wolpert mejoró la idea de los morfógenos de Turing. Propuso el "modelo de la bandera francesa" para explicar cómo los morfógenos actúan en los tejidos en desarrollo. Estas ideas se confirmaron con el trabajo de Christiane Nüsslein-Volhard, quien descubrió el primer morfógeno real, llamado Bicoid, en la mosca de la fruta Drosophila melanogaster.
Hoy en día, la morfogénesis se usa para entender cómo se forma un organismo completo, desde la estructura de cada célula, la capacidad de las células para crear tejidos (diferenciación celular) y cómo se organiza el cuerpo de cada ser vivo (segmentación).
Mecanismos Moleculares de la Morfogénesis
Durante la morfogénesis, varias moléculas son muy importantes. Los morfógenos son moléculas solubles que actúan como señales, controlando la especialización de las células según su concentración. Otro grupo clave son los factores de transcripción, que interactúan con el ADN para decidir qué tipo de célula será cada una. También hay moléculas que controlan cómo las células se mueven (migración) y cómo se pegan entre sí (adhesión).
Por ejemplo, durante la gastrulación (una etapa temprana del desarrollo embrionario), grupos de células madre se sueltan, se mueven a nuevas posiciones dentro del embrión y luego se vuelven a unir para formar nuevos tejidos y órganos.
Mecanismos Celulares de la Morfogénesis
A nivel de los tejidos, la morfogénesis se ve como un aumento de células y su movimiento, lo que cambia la estructura y las interacciones entre ellas. Estos cambios pueden hacer que un tejido se alargue, se haga más delgado, se pliegue, invada otras áreas o se separe en diferentes capas.
Adhesión Célula-Célula

Durante el desarrollo de un embrión, las células se mantienen en capas específicas porque tienen diferentes afinidades para unirse. Esto ocurre cuando las células comparten las mismas moléculas que les permiten adherirse entre sí.
Existen varios tipos de moléculas de adhesión celular (CAM). Un grupo muy importante son las cadherinas. Hay diferentes tipos de cadherinas, y cada una se une preferentemente a su mismo tipo. Por ejemplo, las E-cadherinas (de células epiteliales) se unen a otras E-cadherinas, pero no a las N-cadherinas (de células mesenquimales).
Estas uniones específicas ayudan a mantener los límites entre los grupos de células en un tejido en desarrollo. Además, las células pueden organizarse en capas según la cantidad de moléculas de adhesión que tengan. Las células con mayor adhesión tienden a moverse hacia el centro de una mezcla, mientras que las menos adheridas se quedan en las zonas exteriores.
Experimento de Townes y Holtfreter

La relación entre la adhesión celular y la morfogénesis fue demostrada por Johannes Townes y Philip L. Holtfreter en 1955. Ellos descubrieron que los tejidos de anfibios se separan en células individuales si se colocan en soluciones especiales. Tomaron células de diferentes capas de embriones de anfibios y las mezclaron. Cuando las condiciones volvieron a la normalidad, las células se unieron formando grupos.
Usando embriones de rana y salamandra, Townes y Holtfreter observaron cómo se comportaban las células mezcladas. Los resultados fueron sorprendentes: las células se separaron y se organizaron espacialmente. Por ejemplo, si mezclaban células ectodérmicas y mesodérmicas, las células ectodérmicas se movían hacia el interior. Las células nunca se quedaban mezcladas; siempre se separaban, y a menudo un tipo de tejido envolvía completamente a otro.
Además, los investigadores encontraron que la posición final de las células reagrupadas reflejaba su posición original en el embrión. El ectodermo (capa externa) quedaba en la periferia, el endodermo (capa interna) en el centro, y el mesodermo (capa intermedia) entre ellos. Holtfreter explicó esto como una "afinidad selectiva" entre las células.
Hipótesis de Adhesión Diferencial
Malcolm Steinberg propuso en 1964 la hipótesis de la adhesión diferencial. Esta idea sugiere que las diferencias en la capacidad de las células para adherirse son las responsables de cómo se organizan en estructuras más grandes. Algunas células se "clasifican" a sí mismas, interactuando y agrupándose para maximizar su contacto con células similares, lo que reduce su energía y las hace más estables. Hoy sabemos que las células tienen propiedades tanto de adhesión como de repulsión.
Matriz Extracelular
La morfogénesis celular ocurre en un ambiente lleno de biomoléculas, proteínas y líquidos, conocido como la matriz extracelular (ECM). Los componentes principales son el colágeno, la laminina y la fibronectina, que se organizan en láminas, fibras y geles. Esta matriz actúa como un soporte externo, permitiendo que las células se muevan o cambien de forma según sus programas de especialización.

Las señales de la matriz son recibidas por receptores en la superficie de las células llamados integrinas. Las integrinas pueden reconocer la fibronectina o la laminina. Dentro de la célula, las integrinas se unen a partes del citoesqueleto, como la actina. Estas uniones permiten que las células se muevan sobre la matriz o cambien su forma.
Si la matriz extracelular se altera, el desarrollo normal de algunos órganos puede verse afectado. Por ejemplo, si se cambia la matriz en las glándulas mamarias, su ramificación no se forma correctamente. Otro ejemplo es la pérdida de la unión del corazón en el pez cebra si se elimina la fibronectina. La ausencia de fibronectina también afecta el desarrollo de los riñones.
Citoesqueleto
Los cambios de forma en las células son posibles gracias al citoesqueleto, una red de estructuras internas formada por filamentos de actina, microtúbulos y filamentos intermedios. Los filamentos de actina son delgados y flexibles, y pueden contraerse y relajarse con la ayuda de la miosina para mantener o cambiar la forma de la célula y generar fuerza. Este movimiento se realiza sobre la matriz extracelular. Cuando la miosina se une al ATP, puede moverse y unirse a los filamentos de actina, acercándolos y haciendo que la célula se contraiga.
Además, hay señales externas que pueden regular el citoesqueleto a través de un grupo de proteínas llamadas Rho-Rho, Rac y Cdc42. La activación de estas proteínas permite que los filamentos de actina se formen y que la miosina se active. Rho facilita la contracción de los filamentos. Señales como fuerzas, presión o cambios en la adhesión pueden activar Rho, que a su vez es activado por integrinas y otros receptores. De esta manera, el citoesqueleto recibe señales continuamente. Por ejemplo, un anillo de filamentos de actina y miosina se forma rápidamente para cerrar una herida.

Transición Epitelio-Mesénquima
Uno de los fenómenos más importantes en el desarrollo es la transición epitelio-mesénquima. Es un proceso en el que las células con características epiteliales (que están muy unidas y en contacto con una base) se transforman en células mesenquimales, que son móviles y se separan.
Para que esto ocurra, generalmente disminuyen las uniones mediadas por cadherinas, el citoesqueleto se reorganiza y la matriz extracelular se degrada.
Este proceso es fundamental en varias etapas del desarrollo, como la formación de la cresta neural, la formación del mesodermo en aves o la formación de somitas. También es importante en procesos que ocurren después del desarrollo embrionario, como la curación de heridas o la forma en que algunas células se extienden en enfermedades.
Véase también
 En inglés: Morphogenesis Facts for Kids
En inglés: Morphogenesis Facts for Kids
- Morfógeno
- Diferenciación celular
- Ontogenia
- Organogénesis
- Segmentación (biología)
- Embriogenesis
- Neurulación
- Gastrulación
- Embriogénesis en Drosophila
- Biología del desarrollo
Galería de imágenes
-
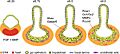
Morfogénesis de una yema hepática. Ocurren cambios en la adhesión celular, modificación de la matriz y transiciones epitelio-mesénquima.

