
Cerebelo para niños
Datos para niños Cerebelo |
||
|---|---|---|
 Encéfalo humano, con el cerebelo marcado en púrpura
|
||
 Cerebelo en púrpura. Sección sagital de un encéfalo en una RMN.
|
||
| Latín | [TA]: cerebellum | |
| TA | A14.1.07.001 | |
| Es parte de | Encéfalo | |
| Arterias | cerebelosa superior, cerebelosa inferior anterior, cerebelosa inferior posterior | |
| Venas | superior del vermis, inferior del vermis, cerebelosas superiores, cerebelosas inferiores, cerebelosa precentral, petrosa | |
| Enlaces externos | ||
| Gray | pág.788 | |
| NeuroLex ID | Cerebellum | |
El cerebelo es una parte muy importante de tu encéfalo. Su función principal es combinar la información que recibes de tus sentidos con las órdenes que tu cerebro envía a tus músculos. Imagina que es como un director de orquesta que se asegura de que todos los movimientos de tu cuerpo sean suaves y precisos.
El cerebelo está conectado con muchas otras partes del encéfalo y con la médula espinal a través de una gran red de nervios. Gracias a esto, puede ajustar y controlar los movimientos que tu corteza cerebral (la parte más grande del cerebro) le pide a tus músculos. También ayuda a mantener tu equilibrio y tu postura.
Si el cerebelo se daña, no sueles quedar paralizado. En cambio, puedes tener problemas para hacer movimientos precisos, mantener el equilibrio o aprender nuevas habilidades motoras. Los primeros estudios en el siglo XVIII ya notaron que las personas con daño en el cerebelo tenían dificultades para coordinar sus movimientos. Investigaciones más recientes han descubierto que el cerebelo también participa en funciones como la atención, el procesamiento del lenguaje, la música y el aprendizaje.
Fue descrito por primera vez por un médico llamado Herófilo hace muchísimos años, en el siglo IV antes de Cristo.

Contenido
- Cerebelo: El Coordinador de tu Cuerpo
- ¿Qué es el Cerebelo y Para Qué Sirve?
- ¿Cómo se Desarrolla el Cerebelo?
- ¿Cómo es el Cerebelo por Fuera?
- ¿Cómo se Divide el Cerebelo?
- ¿Cómo es el Cerebelo por Dentro?
- ¿Cómo se Conecta el Cerebelo con el Resto del Cuerpo?
- ¿Cómo Funcionan las Neuronas del Cerebelo?
- ¿Qué Pasa si el Cerebelo se Daña?
- Galería de imágenes
- Véase también
Cerebelo: El Coordinador de tu Cuerpo
¿Qué es el Cerebelo y Para Qué Sirve?
El cerebelo es un órgano único y central, ubicado en la parte de atrás de tu cráneo. Se encuentra detrás del tronco del encéfalo y debajo de los lóbulos occipitales (la parte de tu cerebro que procesa la vista).
¿Cómo se Desarrolla el Cerebelo?

Como el resto de tu sistema nervioso central y tu piel, el cerebelo se forma a partir de una capa llamada ectodermo durante el desarrollo embrionario.
Al principio, el tubo neural (que luego se convertirá en tu cerebro y médula espinal) tiene tres partes. La parte más baja se llama rombencéfalo. Cuando el embrión tiene unas 5 semanas, el rombencéfalo se divide en dos: el metencéfalo y el mielencéfalo. El metencéfalo es la parte que dará origen al puente troncoencefálico y al cerebelo.
Origen de las Células Cerebelosas
Aunque se pensaba que el cerebelo adulto venía solo del metencéfalo, estudios más recientes han demostrado que sus células también provienen de la parte baja del mesencéfalo. Esto se descubrió usando experimentos con animales, donde se podían seguir el origen de las células.
Control Genético del Desarrollo
El desarrollo del cerebelo está controlado por muchos genes. Por ejemplo, los genes llamados Engrailed (En-1 y En-2) son muy importantes. Si hay mutaciones en el gen En-1, el cerebelo casi no se desarrolla. Si la mutación es en En-2, los daños son menos graves, afectando la forma de los pliegues del cerebelo. Esto sugiere que En-1 define el "territorio" del cerebelo al principio, y En-2 es necesario más tarde.
Etapas de Desarrollo de las Células Granulares
Las células granulares son un tipo de neuronas en el cerebelo. Su desarrollo pasa por cuatro etapas principales:
- Neurogénesis: Cuando se forman nuevas neuronas.
- Diferenciación: Cuando las neuronas empiezan a especializarse.
- Crecimiento axonal y migración: Cuando las neuronas extienden sus "brazos" (axones) y se mueven a su lugar definitivo.
- Formación de conexiones sinápticas: Cuando las neuronas se conectan con otras para comunicarse.
Proliferación de Células Granulares
Las células granulares se multiplican mucho en las primeras semanas después del nacimiento. Una molécula llamada Shh (Sonic hedgehog) es clave para que estas células se dividan. Si se bloquea la acción de Shh, el número de células granulares disminuye mucho.
Diferenciación de Células Granulares
Después de multiplicarse, las neuronas granulares deben dejar de dividirse y empezar a especializarse. Hay genes y proteínas que ayudan a que este proceso ocurra correctamente.
Migración de Células Granulares
Cuando las neuronas granulares inmaduras empiezan a diferenciarse, forman un axón con forma de "T". Luego, se mueven desde su lugar de origen hasta su posición final. Este movimiento es posible gracias a la interacción con otras células de soporte llamadas células gliales de Bergmann. Una proteína llamada astrotactina (ASTN1) es esencial para que estas células se adhieran y puedan migrar.
Formación de Conexiones Sinápticas


Una vez que las neuronas granulares llegan a su destino, se preparan para conectarse con otras neuronas. Sus axones en forma de "T" se conectan con las dendritas de las células de Purkinje. También se forman conexiones especiales llamadas glomérulos sinápticos.
¿Cómo es el Cerebelo por Fuera?
El cerebelo tiene forma de cono aplanado y se divide en dos partes llamadas hemisferios, separadas por una parte central llamada vermis. Tiene tres caras: superior, inferior y anterior.
Cara Superior
La cara superior es como un techo con dos lados inclinados. En el centro, tiene una elevación alargada que se llama vermis superior. A los lados del vermis superior están las caras superiores de los hemisferios cerebelosos. Una fisura profunda, llamada fisura prima, divide esta cara.
Cara Inferior
La cara inferior se apoya sobre una membrana. En el centro, tiene un surco grande llamado vallécula, donde se encuentra el vermis inferior. A los lados, están las caras inferiores de los hemisferios. En la parte de adelante, hay unas protuberancias ovaladas llamadas amígdala cerebelosa, que están cerca del bulbo raquídeo.
Cara Anterior
La cara anterior está muy cerca de la parte de atrás del tronco del encéfalo. Para verla, hay que cortar los tres pares de "tallos" (pedúnculos) que la unen. Tiene una depresión central que forma el techo del cuarto ventrículo (un espacio lleno de líquido en el cerebro).
¿Cómo se Divide el Cerebelo?
El cerebelo se puede dividir de varias maneras: por su forma, por su evolución (filogenética) y por su función.
Divisiones Morfológicas

Esta división describe la superficie del cerebelo. Tiene muchas fisuras (surcos) transversales. Dos de las más profundas, la fisura prima y la fisura posterolateral, lo dividen en tres lóbulos principales:
- Lóbulo anterior: Se encuentra delante de la fisura prima.
- Lóbulo posterior: Se encuentra entre la fisura prima y la posterolateral.
- Lóbulo floculonodular: Se encuentra delante de la fisura posterolateral y está formado por el nódulo y los flóculos.
Divisiones Filogenéticas (Evolutivas)
Desde el punto de vista de la evolución, el cerebelo se divide en tres partes, cada una con una función específica:
- Arquicerebelo: Es la parte más antigua. Se corresponde con el lóbulo floculonodular y está relacionado con el sistema vestibular (del oído interno), que controla el equilibrio.
- Paleocerebelo: Es más moderno que el arquicerebelo. Recibe información de la médula espinal y ayuda a controlar la postura.
- Neocerebelo: Es la parte más nueva y grande. Recibe información de la corteza cerebral y es clave para la coordinación muscular de los movimientos voluntarios.
Representación del Cuerpo en el Cerebelo
Al igual que otras partes del cerebro, el cerebelo tiene un "mapa" de tu cuerpo. El tronco y el cuello, así como las partes cercanas de tus brazos y piernas, están representados en la región del vermis. Las partes de la cara y las partes más lejanas de tus extremidades se localizan en otras zonas.
¿Cómo es el Cerebelo por Dentro?
El cerebelo, como el cerebro, tiene sustancia gris y sustancia blanca. La sustancia gris está en la superficie, formando la corteza cerebelosa, y también en el interior, formando los núcleos profundos. La sustancia blanca se encuentra en el centro, rodeando los núcleos.
La Corteza Cerebelosa
La corteza cerebelosa tiene una superficie muy grande (unos 500 cm²) gracias a sus muchos pliegues. Estos pliegues le dan un aspecto rugoso. Está formada por muchas unidades pequeñas llamadas laminillas cerebelosas.
La sustancia gris de la corteza cerebelosa tiene un grosor de aproximadamente 1 milímetro y está organizada en tres capas con siete tipos principales de neuronas.
Capas de la Corteza

De adentro hacia afuera, las capas son:
- Capa granular: La más profunda, llena de pequeñas neuronas llamadas granos.
- Capa de las células de Purkinje: Formada por los cuerpos de las células de Purkinje, que se organizan en una sola fila.
- Capa molecular: La más superficial, con muchas prolongaciones de células y pocos cuerpos neuronales.
Tipos de Neuronas

Las neuronas de la corteza cerebelosa se dividen en:
- Células de Purkinje: Son las neuronas principales. Son grandes y tienen un árbol dendrítico (como ramas de un árbol) muy ramificado que se extiende por la capa molecular. Sus axones (las "salidas" de la neurona) van hacia los núcleos profundos del cerebelo. Se calcula que hay unos 30 millones de estas neuronas en el cerebelo humano.
- Neuronas intrínsecas (interneuronas): Son neuronas más pequeñas que se quedan dentro de la corteza. Incluyen las células granulares, las células de Golgi, las células de Lugaro, las células estrelladas y las células en cesta. Las células granulares son las más pequeñas del sistema nervioso humano y son muy numerosas (unos 50 mil millones). Las células en cesta son famosas porque sus axones forman una especie de "cesta" alrededor de los cuerpos de las células de Purkinje.
Fibras Nerviosas que Llegan al Cerebelo
Las fibras extrínsecas son los nervios que llegan al cerebelo desde otras partes del sistema nervioso. Las más importantes son:
- Fibras musgosas: Vienen de muchas áreas, como la médula espinal y el tronco del encéfalo. Llevan información de casi todo el sistema nervioso.
- Fibras trepadoras: Vienen de un núcleo llamado olivar inferior. Son muy importantes porque cada una se conecta con varias células de Purkinje y las estimula directamente.
Células de Soporte (Glia)
Además de las neuronas, el cerebelo tiene astrocitos, que son células de soporte. Un tipo especial es la glía de Bergmann, que ayuda a las neuronas a organizarse.
Núcleos Profundos del Cerebelo

Dentro de la sustancia blanca, hay cuatro pares de núcleos de sustancia gris: el núcleo del fastigio, el globoso, el emboliforme y el dentado. El núcleo dentado es el más grande y tiene unas 250.000 neuronas. Estos núcleos reciben información de las células de Purkinje y envían señales a otras partes del cerebro.
Sustancia Blanca
La sustancia blanca del cerebelo tiene una forma que parece un árbol, por eso a veces se le llama "árbol de la vida" (arbor vitae). Está formada por axones (las "salidas" de las neuronas) que conectan diferentes áreas del cerebelo entre sí y con otras partes del sistema nervioso.
¿Cómo se Conecta el Cerebelo con el Resto del Cuerpo?

El cerebelo recibe información de todas las vías motoras y sensitivas de tu cuerpo, y envía señales para controlar tus movimientos. Las señales que salen del cerebelo no suelen ir directamente a los músculos, sino que actúan sobre otros núcleos en el tronco del encéfalo.
Pedúnculos Cerebelosos

El cerebelo se une al tronco del encéfalo mediante tres pares de "tallos" o pedúnculos. Por ellos pasan todas las fibras nerviosas que entran y salen del cerebelo.
Pedúnculos Cerebelosos Inferiores
Conectan el cerebelo con la parte superior del bulbo raquídeo. Por aquí entran fibras que llevan información sobre la posición del cuerpo y el equilibrio, y salen fibras que regulan el equilibrio y los movimientos de los ojos.
Pedúnculos Cerebelosos Medios

Son los más grandes y conectan el cerebelo con el puente troncoencefálico. Por ellos entran muchas fibras que traen información de la corteza cerebral para coordinar los movimientos.
Pedúnculos Cerebelosos Superiores

Conectan el cerebelo con el mesencéfalo. Por aquí entran fibras que llevan información de la médula espinal, la vista y el oído, y salen fibras que controlan los movimientos de los ojos y la coordinación.
Conexiones del Vestibulocerebelo (Equilibrio)
Esta parte del cerebelo se encarga principalmente del equilibrio.
Fibras que Llegan (Aferencias)
Recibe información del sistema vestibular (en el oído interno), que le dice al cerebro dónde está tu cabeza y cómo se mueve tu cuerpo.
Fibras que Salen (Eferencias)
Envía señales a los núcleos vestibulares para regular el equilibrio y a los núcleos que controlan los movimientos de los ojos.
Conexiones del Espinocerebelo (Postura y Movimiento)
Esta parte del cerebelo se encarga de la postura y la coordinación de los movimientos.
Fibras que Llegan (Aferencias)
Recibe información muy rápida de la médula espinal sobre los cambios en tus músculos y extremidades. También recibe información del bulbo raquídeo y del mesencéfalo.
Fibras que Salen (Eferencias)
Envía señales a diferentes núcleos en el tronco del encéfalo y al tálamo para controlar las vías motoras que bajan a la médula espinal.
Conexiones del Cerebrocerebelo (Movimientos Precisos)
Esta es la parte más grande y moderna del cerebelo, crucial para los movimientos voluntarios y precisos.
Fibras que Llegan (Aferencias)
Recibe información de una gran parte de la corteza cerebral (lóbulos frontal, parietal, occipital y temporal). Esta información pasa primero por los núcleos del puente antes de llegar al cerebelo.
Fibras que Salen (Eferencias)
La mayoría de las señales de esta parte del cerebelo salen del núcleo dentado y van al tálamo. Desde el tálamo, las señales regresan a las mismas áreas de la corteza cerebral de donde vinieron.
Conexiones de Sistemas Moduladores
El cerebelo también recibe fibras de sistemas que modulan la actividad cerebral, como los sistemas noradrenérgico y serotoninérgico, que influyen en el estado de ánimo y la atención.
¿Cómo Funcionan las Neuronas del Cerebelo?
Las conexiones neuronales en el cerebelo se pueden ver como un sistema de dos arcos: uno principal que excita y otro secundario que inhibe, y que regula al primero. Este sistema se repite millones de veces en todo el cerebelo.
El Circuito Principal (Excitador)
Este circuito está formado por las fibras musgosas y trepadoras que llegan a los núcleos profundos del cerebelo. Las neuronas de estos núcleos envían señales excitadoras a otras partes del cerebro.
El Circuito Secundario (Inhibidor)
Este circuito pasa por la corteza cerebelosa y su pieza clave es la célula de Purkinje. Las células de Purkinje reciben señales excitadoras e inhibidoras. Sus axones envían señales inhibidoras a los núcleos profundos, modulando así el circuito principal.
Circuitos Excitadores de las Células de Purkinje
Las células de Purkinje pueden ser estimuladas de dos maneras:
- Directamente: Por las fibras trepadoras, que producen una estimulación muy fuerte y específica.
- Indirectamente: Por las fibras musgosas, a través de las células granulares.
Circuitos Inhibidores de las Células de Purkinje
Las interneuronas inhibitorias (células de Golgi, estrelladas y en cesta) modulan la actividad de las células de Purkinje. Por ejemplo, las células estrelladas y en cesta actúan directamente sobre las células de Purkinje, mientras que las células de Golgi actúan indirectamente a través de las células granulares.
Aprendizaje Motor
El cerebelo es fundamental para el aprendizaje motor, es decir, para aprender y perfeccionar habilidades como andar en bicicleta, tocar un instrumento o practicar un deporte.
¿Qué Pasa si el Cerebelo se Daña?
Cuando el cerebelo sufre una lesión, los síntomas suelen ser:
Síntomas Comunes
- Hipotonía: Los músculos se sienten más blandos de lo normal y tienen menos resistencia.
- Ataxia o descoordinación de los movimientos voluntarios: Dificultad para coordinar movimientos precisos. Por ejemplo, al intentar tocarse la nariz con el dedo, la persona puede pasarse o no llegar. También hay dificultad para hacer movimientos rápidos y alternados.
- Alteración del equilibrio y de la marcha: La persona se siente inestable al estar de pie y al caminar, y puede tambalearse como si estuviera mareada. La marcha es característica, como la de alguien que ha bebido demasiado.
- Temblor intencional: Un temblor que aparece o se hace más evidente al intentar hacer un movimiento.
Síndrome del Vermis Cerebeloso
Si la lesión afecta el vermis (la parte central del cerebelo), los problemas de coordinación afectan principalmente la cabeza y el tronco. La persona puede tener dificultad para mantener la cabeza quieta o el tronco erguido, y tiende a caerse hacia adelante o hacia atrás.
Síndrome Hemisférico Cerebeloso
Si la lesión está en uno de los hemisferios del cerebelo, los síntomas suelen ser en el mismo lado del cuerpo que la lesión. Los movimientos de los brazos y piernas se ven afectados, con mucha descoordinación. También puede haber dificultad para hablar (disartria) y movimientos involuntarios de los ojos (nistagmo).
Causas de Problemas en el Cerebelo
Las causas más comunes de problemas en el cerebelo incluyen:
- Problemas vasculares: Como infartos o hemorragias.
- Tumores: Como el meduloblastoma (común en niños) o el astrocitoma.
- Traumatismos: Golpes o lesiones en la cabeza.
- Infecciones: Como cerebelitis (inflamación del cerebelo).
- Enfermedades degenerativas: Que causan un deterioro progresivo del cerebelo.
- Problemas autoinmunes: Cuando el sistema de defensa del cuerpo ataca por error al cerebelo.
- Malformaciones: Problemas en el desarrollo del cerebelo desde el nacimiento.
Galería de imágenes
-
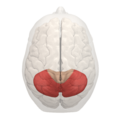
Lóbulo posterior en rojo
-
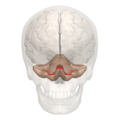
Lóbulo floculonodular
-
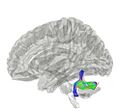
Pedúnculos cerebelosos inferiores
-
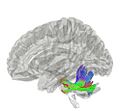
Pedúnculos cerebelosos medios
-
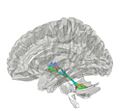
Pedúnculos cerebelosos superiores
-
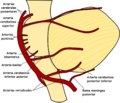
Representación esquemática de las arterias del cerebelo y del tronco encefálico



{kind=link}
Véase también
 En inglés: Cerebellum Facts for Kids
En inglés: Cerebellum Facts for Kids
- Cerebro
- Encéfalo
- Sistema nervioso central
- Nervio
- Trastornos neurológicos relacionados con el gluten
