
Sustancia gris para niños
Datos para niños Sustancia gris |
||
|---|---|---|
 Formación del nervio espinal a partir de las raíces dorsal y ventral. (Sustancia gris etiquetada en el centro a la derecha).
|
||
| Nombre y clasificación | ||
| Latín | [TA]: substantia grisea | |
| TA | A14.1.00.002 | |
|
|
||
La sustancia gris (también conocida como materia gris) es una parte muy importante de tu sistema nervioso central. Tiene un color grisáceo y está formada principalmente por las partes principales de las neuronas, llamadas somas, y por sus "cables" (llamados axones) que no tienen una cubierta especial llamada mielina. También contiene otras células de apoyo llamadas gliales.
Contenido
La Sustancia Gris: El Centro de Pensamiento de tu Cerebro
¿Qué es la Sustancia Gris y Dónde se Encuentra?
La sustancia gris es fundamental para el funcionamiento de tu cerebro y médula espinal. En la médula espinal, la puedes ver en el centro, con una forma que parece una mariposa o la letra H. En cambio, en el cerebro, la sustancia gris se encuentra en la parte exterior, formando la corteza cerebral. Esta corteza es la parte más compleja de todo tu sistema nervioso.
También hay grupos de sustancia gris en el interior del cerebro, como los ganglios basales. Estos actúan como estaciones de relevo, ayudando a que la información se mueva correctamente.
¿Qué Hace la Sustancia Gris?
A diferencia de otras partes del cerebro que tienen mielina y transmiten información muy rápido, la sustancia gris no tiene esta cubierta. Esto significa que no transmite los impulsos nerviosos tan velozmente. Sin embargo, esta característica es clave para su función principal: el procesamiento de información. Es decir, la sustancia gris es donde ocurre el razonamiento, el pensamiento y la toma de decisiones.
Se ha pensado que la cantidad de sustancia gris podría estar relacionada con la inteligencia de un ser vivo. Pero esto no es una regla estricta. Por ejemplo, los delfines tienen más sustancia gris que los humanos. Esto les da habilidades cerebrales diferentes a las nuestras.
La Sustancia Gris en la Médula Espinal

En la médula espinal, la sustancia gris tiene una forma de "H" o mariposa. Esta "H" tiene diferentes partes:
- Asta dorsal: Esta parte recibe información de los sentidos, como el tacto o el dolor.
- Asta intermediolateral: Se encuentra solo en algunas secciones de la médula. Contiene neuronas que controlan funciones automáticas del cuerpo.
- Asta ventral: Esta parte envía señales a los músculos para que se muevan.
- Zona intermedia: Contiene neuronas que conectan diferentes partes de la médula.
Tipos de Neuronas en la Sustancia Gris
Dentro de la sustancia gris, hay diferentes tipos de neuronas que cumplen funciones específicas:
- Neuronas radiculares: Se encuentran en la parte delantera de la médula. Sus "cables" (axones) salen directamente del sistema nervioso central para controlar los músculos. Hay dos tipos principales:
* Motoneuronas alfa: Son grandes y controlan los movimientos de tus músculos. * Motoneuronas gamma: Son más pequeñas y también ayudan a controlar los músculos, especialmente en los receptores que detectan el estiramiento muscular.
- Neuronas cordonales: Estas neuronas conectan diferentes partes de la médula espinal, tanto hacia arriba y abajo como de un lado a otro. Algunas de ellas llevan información sensorial.
- Interneuronas (o neuronas Golgi tipo II): Son neuronas pequeñas que conectan otras neuronas cercanas dentro de la sustancia gris. Ayudan a coordinar la actividad neuronal.
¿Cómo Funcionan los Reflejos?
La médula espinal no es solo un camino para la información. También puede organizar respuestas por sí misma, como los reflejos. Un reflejo es una respuesta rápida e involuntaria a un estímulo. Por ejemplo, cuando el médico te golpea suavemente la rodilla y tu pierna se estira, eso es un reflejo.
Los reflejos son muy importantes para mantener el equilibrio y la postura de tu cuerpo. La base de un reflejo es el "arco reflejo", que incluye:
- Un órgano que detecta el estímulo (receptor).
- Una neurona aferente que lleva la señal al sistema nervioso.
- Una neurona eferente que lleva la señal de respuesta.
- Un órgano que realiza la acción (efector), como un músculo.
Algunos reflejos son muy rápidos porque solo involucran una conexión (sinapsis) entre neuronas. Los reflejos nos permiten evaluar cómo funcionan diferentes partes de la médula espinal.
La sustancia gris es muy sensible a la falta de oxígeno y a ciertas condiciones. Por eso, si los reflejos no funcionan bien, puede ser una señal importante para los médicos.
Galería de imágenes
-
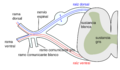
Formación del nervio espinal a partir de las raíces dorsal y ventral. (Sustancia gris etiquetada en el centro a la derecha).
-
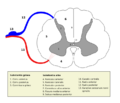
Astas ventral y dorsal, y la zona intermedia señaladas con el número 1, 2 y 3, respectivamente
Véase también
 En inglés: Grey matter Facts for Kids
En inglés: Grey matter Facts for Kids
