
Historia de la teoría de la membrana celular para niños
La historia de la teoría celular comenzó con las primeras veces que se vieron células de organismos usando microscopios en el siglo XVII. Para el siglo XIX, los científicos ya pensaban que debía haber una barrera especial alrededor de la célula. Esta barrera separaría el interior de la célula de lo que la rodea.
Al estudiar cómo funcionaban algunas sustancias que adormecen, se pensó que esta barrera, o membrana celular, debía estar hecha de algún tipo de lípido (un tipo de grasa). Pero su estructura exacta no se supo hasta 1925. En ese año, varios experimentos mostraron que la membrana estaba formada por dos capas de moléculas de lípidos, como un sándwich, lo que se llamó una bicapa lipídica.
Durante las décadas siguientes del siglo XX, nuevos experimentos descubrieron que también había proteínas unidas a la membrana celular. Aunque al principio no se ponían de acuerdo sobre qué hacían estas proteínas. En los años 70, surgió el modelo de mosaico fluido. Este modelo dice que las proteínas "flotan" libremente en la bicapa lipídica, atravesando la membrana por la mitad o por completo. Este modelo, aunque es una forma sencilla de verlo, se ha usado mucho desde entonces.

Contenido
Primeras ideas sobre la barrera celular
Desde que se inventó el microscopio en el siglo XVII, se supo que los tejidos de plantas y animales están hechos de células. El científico Robert Hooke fue quien usó por primera vez la palabra "células" para describirlas.
La pared celular de las plantas se veía muy bien incluso con los primeros microscopios. Pero en las células animales no se veía una estructura parecida, aunque se creía que debía existir algo. A mediados del siglo XIX, muchos científicos investigaban esto. Moritz Traube sugirió que la capa exterior de las células animales debía ser semipermeable. Esto significaba que permitía el paso de algunas partículas, como los iones (partículas con carga eléctrica). Traube no tenía pruebas directas de qué estaba hecha esta capa.
El primer investigador que pensó que la membrana celular era de tipo lipídico (graso) fue Quincke. Él notó que una célula suele tener forma de esfera en el agua. Y si se partía por la mitad, formaba dos esferas más pequeñas, igual que el aceite. Quincke también se dio cuenta de que una capa fina de aceite se comporta como una membrana semipermeable. Por eso, Quincke concluyó que la membrana celular estaba hecha de una capa de grasa líquida muy fina.
Más tarde, Hans Meyer y Ernest Overton observaron que las moléculas con efectos adormecedores suelen disolverse tanto en agua como en aceite. Pensaron que, para pasar por la membrana celular, una molécula debía ser al menos un poco soluble en aceite. A esto lo llamaron la "teoría lipídica de los efectos adormecedores". Con esta idea y otros experimentos, concluyeron que la membrana celular estaba hecha de lecitina y colesterol. Una de las primeras críticas a esta teoría fue que no explicaba cómo la célula elegía qué sustancias dejar pasar. Este problema no se resolvió hasta casi 50 años después. Fue cuando se descubrieron unas moléculas especiales llamadas proteínas integrales de membrana que podían actuar como "bombas" para transportar iones.
Descubriendo la estructura de la bicapa lipídica

A principios del siglo XX, se sabía de qué estaba hecha la membrana celular, pero no cómo estaba organizada. Dos experimentos en 1942 intentaron resolver esto. En uno, Hugo Fricke midió el grosor de la membrana celular de los eritrocitos (glóbulos rojos). Aunque su experimento fue preciso, no midió el grosor completo de la membrana. Por eso, concluyó por error que la membrana tenía solo una capa de lípidos.
En otro experimento, Gorter y Grendel lo vieron de otra manera. Extrajeron los lípidos de los glóbulos rojos y los extendieron en una capa muy fina. Al comparar el área de esta capa con el área de las células, encontraron que la capa era el doble de grande. Aunque más tarde se vieron algunos problemas en este experimento, la conclusión de que la membrana estaba formada por una bicapa lipídica (dos capas) fue correcta.
Una década después, Davson y Danielli propusieron que la bicapa lipídica estaba cubierta por ambos lados con una capa de proteínas. Este modelo resultó ser incorrecto.
El uso del microscopio electrónico a finales de los años 50 ayudó a investigar la estructura de la membrana de forma más directa. J. David Robertson propuso el concepto de "unidad de membrana". Él dijo que la estructura de bicapa era común a todas las membranas celulares y también a las membranas de los organelos (partes internas de la célula).
Cómo evolucionó la teoría de la membrana
Las ideas de una membrana semipermeable y de una barrera que dejaba pasar algunos líquidos pero no otras moléculas se desarrollaron al mismo tiempo. La palabra ósmosis (el paso de agua a través de una membrana) se usó desde 1827. En 1877, el botánico Wilhelm Friedrich Philipp Pfeffer propuso que la célula estaba envuelta por una superficie delgada, la membrana plasmática.
Overton fue el primero en proponer la idea de una membrana plasmática hecha de lípidos en 1899.
La mayor debilidad de la teoría de la membrana lipídica era que no explicaba por qué el agua pasaba tan fácilmente. Para resolver esto, Nathansohn propuso en 1904 la teoría del mosaico. Según esta, la membrana era como un mosaico con zonas de lípidos y zonas con un gel semipermeable. Ruhland mejoró esta teoría añadiendo poros que permitían el paso de moléculas pequeñas.
En 1939, Harvey y Danielli presentaron un modelo de membrana con una bicapa lipídica cubierta por proteínas en cada lado. Esto ayudaba a explicar algunas mediciones.
El concepto de la bomba de membrana
Con el uso de isótopos radiactivos, se demostró que las células no son impermeables al ion sodio (Na+). Esto era difícil de explicar con la teoría de la barrera de membrana. Así surgió la idea de que existía una "bomba de sodio" que sacaba el Na+ de la célula. Este nuevo concepto sugería que las células están en un estado de equilibrio dinámico. Esto significa que usan energía constantemente para mantener las diferencias de concentración de iones dentro y fuera de la célula.
En 1935, Karl Lohmann descubrió el ATP y su papel como fuente de energía para las células. Él introdujo la idea de una bomba de sodio que funcionaba gracias a la energía de la célula. El trabajo de Hodgkin, Huxley y Katz apoyó mucho la idea de la bomba de sodio.
El Modelo de Mosaico Fluido

Mientras se desarrollaba el modelo de la bicapa, Paul Mueller y Donald Rudin lograron crear membranas similares en el laboratorio. Pudieron estudiar sus propiedades y vieron que eran fluidas, tenían alta resistencia eléctrica y podían repararse solas si se pinchaban.
En 1970, Frye y Edidin demostraron de forma clara la misma fluidez en la superficie de las células. Fusionaron dos células con diferentes marcadores de color en su membrana. Vieron cómo las dos superficies se mezclaban al unirse. Los resultados de este experimento fueron clave para que en 1972, Singer y Nicolson desarrollaran el modelo de "mosaico fluido" para la membrana celular.
Según este modelo, las membranas biológicas están hechas de una bicapa lipídica pura. Las proteínas la atraviesan por la mitad o por completo. Estas proteínas flotan libremente en la bicapa líquida. El modelo de mosaico fluido fue el primero en unir todos los elementos: un líquido, canales en la membrana y diferentes formas en que la bicapa y las proteínas se unen, en una sola teoría.
Investigación moderna de la membrana celular
La investigación continua de la membrana celular ha mostrado que el modelo de mosaico fluido es un poco más simple de lo que es en realidad. Por ejemplo, se ha descubierto que las proteínas de canal iónico no son solo tubos de agua. Además, la difusión libre en la superficie celular no ocurre en todas partes. A menudo se limita a áreas pequeñas. Esto se debe a que la membrana está anclada al citoesqueleto (el "esqueleto" de la célula), a la separación de las grasas y a la unión de las proteínas.
Estudios recientes también indican que la parte de la membrana plasmática que es solo lípidos es más pequeña de lo que se pensaba. La mayor parte de la superficie celular está unida a alguna proteína. A pesar de estas nuevas observaciones, el modelo de mosaico fluido sigue siendo popular. Se usa a menudo para dar una idea general de cómo son las membranas biológicas.
Galería de imágenes
-
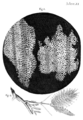
Boceto del corcho visto a través de un microscopio. El corcho fue uno de los primeros objetos examinados por Robert Hooke a través de su microscopio y encontró que estaba compuesto de cientos de pequeños bolsillos que nombró "células".
-
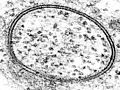
Una micrografía de la transmisión obtenida por un microscopio electrónico mostrando una vesícula de lípidos. Las dos bandas oscuras son los dos prospectos que conforman la bicapa. Imágenes similares tomadas en los cincuenta y sesenta confirmaron la naturaleza de la bicapa en la membrana celular.
-

Diagrama de una membrana celular mostrando proteínas de membrana integrales y periféricas.
Véase también
 En inglés:
En inglés: 