
Glucólisis para niños
| Reacción global de la glucólisis |
|---|
 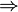  + + |
| α-D-glucosa + 2NAD+ + 2ADP + 2Pi ==> 2(piruvato) + 2NADH + 2ATP + 2H+ + 2H2O |
La glucólisis (que significa "ruptura del azúcar" en griego) es un proceso muy importante que ocurre dentro de nuestras células. Es como una fábrica que toma la glucosa (un tipo de azúcar que obtenemos de los alimentos) y la transforma para conseguir energía. Esta energía es vital para que la célula pueda hacer todas sus funciones.
Este proceso se lleva a cabo en 10 pasos seguidos, cada uno ayudado por una enzima diferente. Al final, una molécula de glucosa se convierte en dos moléculas más pequeñas llamadas piruvato. El piruvato puede seguir otros caminos para generar aún más energía para el cuerpo. La glucólisis puede ocurrir tanto si hay oxígeno presente como si no lo hay. Cuando no hay oxígeno, se le llama proceso anaeróbico.
La forma más conocida de glucólisis es la llamada vía de Embden-Meyerhof, nombrada así por los científicos que la explicaron por primera vez. Es el primer paso para que el cuerpo use los carbohidratos como fuente de energía.
Contenido
¿Cómo se descubrió la glucólisis?
Los primeros indicios de la glucólisis se vieron en 1860, cuando Louis Pasteur descubrió que los microorganismos eran los responsables de la fermentación. Más tarde, en 1897, Eduard Buchner encontró que un extracto de células podía causar fermentación.
En 1905, Arthur Harden y William Young descubrieron que para que la fermentación ocurriera, se necesitaban dos partes: una parte con enzimas (que son como herramientas especiales) y otra parte con moléculas pequeñas como el ATP (la moneda de energía de la célula). Los detalles de cómo funciona la glucólisis se entendieron mejor alrededor de 1940, gracias al trabajo de Otto Meyerhoff y Luis Leloir. Fue difícil descubrir todos los pasos porque las moléculas intermedias duran muy poco tiempo y están en bajas cantidades.
En la mayoría de los seres vivos, desde las bacterias más simples hasta los animales y las plantas, la glucólisis ocurre en el citosol de la célula (que es como el "líquido" dentro de la célula). En las plantas, algunas reacciones de la glucólisis también se encuentran en los cloroplastos, donde ocurre la fotosíntesis. Como esta vía es tan antigua y se encuentra en tantos organismos, se cree que es uno de los procesos metabólicos más antiguos de la vida.
¿Cómo funciona la glucólisis?

Durante la glucólisis, la célula obtiene una ganancia neta de dos moléculas de ATP. El ATP es la principal fuente de energía que la célula usa para realizar sus tareas. También se producen moléculas de NADH, que pueden usarse para otras reacciones o para generar más ATP si hay oxígeno. Si no hay oxígeno, el NADH ayuda a transformar el piruvato en otras sustancias, como el ácido láctico o el etanol, en un proceso llamado fermentación.
La glucólisis es la forma más rápida que tiene una célula para conseguir energía. Por eso, cuando el cuerpo necesita energía de los carbohidratos, la glucólisis es la primera vía que se activa. Este proceso tiene 10 reacciones enzimáticas que transforman una molécula de glucosa en dos moléculas de piruvato.
La glucólisis se divide en dos fases principales:
- La primera fase: Es una fase de "gasto de energía". Aquí, la célula usa 2 moléculas de ATP para preparar la glucosa y dividirla en dos moléculas más pequeñas llamadas gliceraldehído. Esto es como invertir un poco de energía para luego obtener más.
- La segunda fase: Es una fase de "obtención de energía". En esta fase, las dos moléculas de gliceraldehído se transforman en compuestos con mucha energía. Al final de esta fase, se producen 4 moléculas de ATP. Como se gastaron 2 ATP en la primera fase, la ganancia neta es de 2 ATP.
¿Qué pasa después de la glucólisis?
Una vez que la glucosa se ha transformado en dos moléculas de piruvato, el camino que sigue el piruvato depende de si hay oxígeno o no:
- Con oxígeno: En los organismos que usan oxígeno (aeróbicos), el piruvato entra en un proceso llamado ciclo de Krebs. Este ciclo produce más moléculas de energía como NADH y FADH2. Estas moléculas luego pasan por la cadena de transporte de electrones, donde se genera una gran cantidad de ATP. De esta manera, una sola molécula de glucosa puede producir hasta 30 moléculas de ATP.
- Sin oxígeno: Cuando las células no tienen suficiente oxígeno (como en los músculos durante un ejercicio intenso o en algunas bacterias), el piruvato se transforma mediante la fermentación. Este proceso es menos eficiente, ya que solo produce 2 moléculas de ATP por cada molécula de glucosa.
* En las levaduras, la fermentación produce etanol (alcohol) y CO2 (gas). Esto se usa para hacer pan o bebidas. * En los músculos y en los eritrocitos (glóbulos rojos), la fermentación produce ácido láctico. Este ácido es lo que a veces causa la sensación de "quemazón" en los músculos cuando hacemos mucho ejercicio.
¿Para qué sirve la glucólisis?
Las funciones principales de la glucólisis son:
- Producir moléculas de alta energía (ATP y NADH) que la célula usa como combustible. Esto ocurre tanto en la respiración aeróbica (con oxígeno) como en la fermentación (sin oxígeno).
- Generar piruvato, que es una molécula clave que puede entrar al ciclo de Krebs para producir mucha más energía.
- Crear moléculas intermedias de 6 y 3 carbonos que pueden ser usadas en otros procesos importantes de la célula.
Las etapas de la glucólisis
La glucólisis se divide en dos fases principales, cada una con varias reacciones:
Fase de gasto de energía (se usa ATP)
En esta primera parte, la glucosa se prepara y se divide en dos moléculas más pequeñas. Aquí se "invierte" energía en forma de ATP.
- 1.er paso: Hexoquinasa
La glucosa recibe un grupo fosfato de una molécula de ATP, lo que la "activa" y la convierte en glucosa-6-fosfato. Esto es importante porque la glucosa-6-fosfato no puede salir de la célula, asegurando que la energía se quede dentro.
 |
| Glucosa + ATP |
- 2.º paso: Glucosa-6-P isomerasa
La glucosa-6-fosfato cambia su forma para convertirse en fructosa-6-fosfato. Este cambio es importante para los siguientes pasos.
 |
| Glucosa-6-fosfato |
- 3.er paso: Fosfofructoquinasa
La fructosa-6-fosfato recibe otro grupo fosfato de otro ATP, convirtiéndose en fructosa-1,6-bisfosfato. Esta es una reacción clave y es el punto principal de control de la glucólisis.
 |
| Fructosa-6-fosfato + ATP |
- 4.º paso: Aldolasa
La fructosa-1,6-bisfosfato se rompe en dos moléculas de tres carbonos: dihidroxiacetona fosfato y gliceraldehído-3-fosfato.
 |
| Fructosa-1,6-bisfosfato |
- 5.º paso: Triosa fosfato isomerasa
Como solo el gliceraldehído-3-fosfato puede seguir en la glucólisis, la dihidroxiacetona-fosfato se convierte en otra molécula de gliceraldehído-3-fosfato. Así, a partir de una glucosa, ahora tenemos dos moléculas de gliceraldehído-3-fosfato listas para la siguiente fase.
 |
| Dihidroxiacetona-fosfato |
Fase de beneficio energético (se obtiene ATP y NADH)
En esta segunda etapa, las dos moléculas de gliceraldehído-3-fosfato se transforman en piruvato, y en el proceso se genera energía.
- 6.º paso: Gliceraldehído-3-fosfato deshidrogenasa
El gliceraldehído-3-fosfato se oxida y se le añade un fosfato, usando una molécula de NAD+ que se convierte en NADH. Esto aumenta la energía del compuesto.
 |
NAD+ NADH + Pi + H+ Gliceraldehído-3-fosfato deshidrogenasa |
 |
| Gliceraldehído-3-fosfato + Pi + NAD+ |
1,3-Bisfosfoglicerato + NADH + H+ |
- 7.º paso: Fosfoglicerato quinasa
El grupo fosfato de la molécula de alta energía (1,3-bisfosfoglicerato) se transfiere a una molécula de ADP, formando la primera molécula de ATP. Como tenemos dos moléculas de gliceraldehído-3-fosfato, se producen 2 ATP en este paso.
|
ADP ATP Fosfoglicerato quinasa |
 |
| 1,3-Bisfosfoglicerato + ADP |
3-Fosfoglicerato + ATP |
- 8.º paso: Fosfoglicerato mutasa
El fosfato de la molécula de 3-fosfoglicerato cambia de posición, moviéndose del carbono 3 al carbono 2, formando 2-fosfoglicerato.
|
Fosfoglicerato mutasa |
 |
| 3-Fosfoglicerato | 2-Fosfoglicerato |
- 9.º paso: Enolasa
Se elimina una molécula de agua del 2-fosfoglicerato, creando un doble enlace y formando fosfoenolpiruvato, una molécula con mucha energía.
|
enolasa |
 |
| 2-Fosfoglicerato | Fosfoenolpiruvato + H2O |
- 10.º paso: Piruvato quinasa
El fosfato del fosfoenolpiruvato se transfiere a una molécula de ADP, formando la segunda molécula de ATP. Como son dos moléculas de fosfoenolpiruvato, se producen 2 ATP más. El producto final de la glucólisis es el piruvato.
|
ADP ATP piruvato quinasa |
 |
| Fosfoenolpiruvato + ADP | Piruvato + ATP |
En resumen, de una molécula de glucosa, la glucólisis produce una ganancia neta de 2 moléculas de ATP y 2 moléculas de NADH. El piruvato que se forma puede seguir otros caminos para generar más energía, como el ciclo de Krebs si hay oxígeno.
¿Cómo se controla la glucólisis?
La glucólisis es un proceso muy importante y está cuidadosamente regulado por la célula para asegurarse de que se produzca la cantidad correcta de energía en el momento adecuado. Esto se hace principalmente controlando la actividad de algunas enzimas clave.
El efecto Pasteur
Efecto Pasteur Louis Pasteur descubrió que la presencia de oxígeno disminuye la velocidad de la fermentación en las levaduras. Esto significa que las células de levadura prefieren usar oxígeno para obtener energía cuando está disponible, porque es más eficiente. Este "efecto Pasteur" fue una de las primeras pistas de que el metabolismo de la glucosa podía cambiar dependiendo de si había oxígeno o no.
Regulación por enzimas

La glucólisis se controla en tres puntos principales, donde las reacciones son irreversibles:
- La enzima hexoquinasa (en el primer paso) se inhibe si ya hay mucha glucosa-6-fosfato, lo que indica que la célula no necesita más.
- La fosfofructoquinasa-1 (en el tercer paso) es la enzima más importante para regular la glucólisis. Es como una "llave de paso": si está muy activa, se produce mucha energía; si está inhibida, se produce poca. Esta enzima se activa cuando la célula necesita energía (cuando hay mucho ADP o AMP) y se inhibe cuando ya tiene suficiente energía (cuando hay mucho ATP o citrato).
- La piruvatoquinasa (en el último paso) también se regula. En el hígado, por ejemplo, se inhibe si hay mucho ATP, lo que indica que la célula tiene suficiente energía.
Regulación por hormonas
Las hormonas también juegan un papel importante en la regulación de la glucólisis:
- Cuando los niveles de glucosa en la sangre aumentan (después de comer), el páncreas produce insulina. La insulina aumenta la actividad de algunas enzimas de la glucólisis en el hígado, lo que ayuda a que la glucosa entre en las células y se use para energía.
- Cuando los niveles de glucosa en la sangre son bajos, el páncreas produce glucagón. El glucagón disminuye la glucólisis y estimula la producción de nueva glucosa en el hígado, para mantener los niveles de azúcar en la sangre estables.
Producción de glucosa
Así como la glucólisis rompe la glucosa para obtener energía, existe otro proceso llamado gluconeogénesis que hace lo contrario: crea nueva glucosa. Esto es muy importante cuando el cuerpo necesita azúcar pero no la está obteniendo de los alimentos. La gluconeogénesis se realiza principalmente en el hígado y en los riñones, usando otras moléculas como el piruvato o el lactato.
La gluconeogénesis es estimulada por el glucagón (cuando el azúcar en sangre es bajo) y es inhibida por la insulina (cuando el azúcar en sangre es alto).
Glucólisis en plantas
Las plantas son especiales porque pueden producir su propia glucosa a través de la fotosíntesis. Esta glucosa es su fuente de energía. Las plantas también realizan la glucólisis y la respiración para obtener energía de esa glucosa, al igual que los animales. La respiración en las plantas ocurre todo el tiempo, no solo cuando hay luz. Durante la respiración, las plantas toman oxígeno y liberan dióxido de carbono y vapor de agua a través de unas pequeñas aberturas en sus hojas llamadas estomas.
Véase también
 En inglés: Glycolysis Facts for Kids
En inglés: Glycolysis Facts for Kids
- Metabolismo
- Ciclo de Krebs
- Fermentación
- Fosforilación oxidativa
- Vía metabólica
