
Linfocito T CD4+ para niños
Los linfocitos T CD4+ son un tipo especial de linfocitos, que son células muy importantes de nuestro sistema inmunitario. También se les llama linfocitos T colaboradores o linfocitos T de ayuda. Su trabajo principal es ayudar a otras células del sistema inmunitario a defender nuestro cuerpo de enfermedades.
Estas células son como los "directores de orquesta" del sistema inmunitario. No atacan directamente a los gérmenes o a las células infectadas. En cambio, activan y guían a otras células para que hagan ese trabajo. Son esenciales para la inmunidad adaptativa, que es la parte de nuestras defensas que aprende a reconocer y combatir amenazas específicas.
Los linfocitos T colaboradores ayudan a los linfocitos B a producir anticuerpos, que son como "misiles guiados" contra los invasores. También activan a los linfocitos T citotóxicos (que son los "soldados" que destruyen células infectadas) y aumentan la capacidad de los fagocitos, como los macrófagos, para eliminar bacterias. Por eso se les llama "colaboradores", porque su función es ayudar a los demás.
Los linfocitos T colaboradores siempre tienen una proteína en su superficie llamada CD4. Por eso se les conoce como linfocitos T CD4+.
La importancia de estas células se ve claramente en enfermedades como la infección por el VIH. Este virus ataca y destruye los linfocitos T CD4+. Cuando el número de estas células disminuye mucho, el sistema inmunitario se debilita, lo que lleva a una condición conocida como sida (síndrome de inmunodeficiencia adquirida). Esto hace que el cuerpo sea muy vulnerable a otras infecciones. También existen enfermedades genéticas raras que causan problemas con los linfocitos T CD4+, lo que puede tener consecuencias graves para la salud.
Contenido
Activación de los linfocitos T colaboradores

Cuando los linfocitos T colaboradores son jóvenes y aún no han encontrado un antígeno (una sustancia extraña, como parte de un virus o bacteria), se les llama "células T vírgenes". Estas células salen del timo (un órgano donde maduran) y viajan por el cuerpo, llegando a lugares como los ganglios linfáticos.
Los linfocitos T CD4+ tienen un "receptor" especial en su superficie llamado TcR (receptor de células T). Este receptor es como una antena que busca antígenos específicos. También tienen la proteína CD4, que les ayuda a reconocer ciertas moléculas llamadas MHC de clase II.
Las moléculas MHC de clase II solo se encuentran en la superficie de unas células especiales llamadas células presentadoras de antígeno (CPA). Las CPA más importantes son las células dendríticas, los macrófagos y los linfocitos B. Su trabajo es "mostrar" los fragmentos de antígenos a los linfocitos T.
Reconocimiento: La primera señal

Cuando hay una infección, las células presentadoras de antígeno (CPA) "capturan" el material extraño (como bacterias o virus). Luego, lo procesan y viajan a un ganglio linfático. Allí, las CPA muestran pequeños fragmentos del antígeno unidos a sus moléculas MHC de clase II.
Cuando un linfocito T colaborador virgen encuentra una CPA que le presenta un antígeno que reconoce, su receptor TcR y la proteína CD4 se unen fuertemente al complejo antígeno-MHC de la CPA. Esta unión es como la "primera señal" para activar el linfocito T. Es el primer paso para que el linfocito T colaborador sepa que hay una amenaza.
Verificación: La segunda señal

Después de recibir la primera señal, el linfocito T virgen necesita una "segunda señal" para confirmar que el antígeno es realmente una amenaza externa. Esta segunda señal es como una verificación de seguridad. Si no recibe esta segunda señal, la célula T podría pensar que el antígeno es parte de nuestro propio cuerpo y se volvería "anérgica", lo que significa que no responderá a ese antígeno en el futuro.
Esta segunda señal ocurre cuando una proteína llamada CD28 en la superficie del linfocito T se une a otras proteínas (CD80 o CD86) en la CPA. Esta "coestimulación" es crucial para que el linfocito T se active correctamente y no ataque por error a nuestras propias células.
Una vez que ambas señales se han activado, el linfocito T colaborador está listo para actuar. Si el cuerpo se vuelve a encontrar con el mismo antígeno en el futuro, los linfocitos T de memoria (que son células T que "recuerdan" el antígeno) solo necesitarán la primera señal para activarse rápidamente. Esto explica por qué somos inmunes a algunas enfermedades después de haberlas tenido o de habernos vacunado.
Proliferación: Multiplicarse para la defensa
Cuando el linfocito T colaborador recibe ambas señales, empieza a multiplicarse rápidamente. Para lograr esto, libera una sustancia llamada interleucina-2 (IL-2), que es como un "fertilizante" para su propio crecimiento. Los linfocitos T activados también producen un receptor especial para la IL-2, lo que les permite unirse a ella y seguir multiplicándose.
Esta multiplicación es muy importante porque crea un gran número de linfocitos T colaboradores listos para combatir la infección.
Maduración: Especialización de las células T
Después de multiplicarse, los linfocitos T colaboradores se diferencian en varios tipos de células especializadas:
- Linfocitos T efectores: Son las células que actúan de inmediato. Liberan citoquinas (mensajeros químicos) que estimulan a otras células inmunitarias para que luchen contra la infección.
- Linfocitos T de memoria: Estas células "recuerdan" el antígeno. Permanecen en el cuerpo por mucho tiempo y, si el mismo antígeno vuelve a aparecer, se activan rápidamente para dar una respuesta inmune más fuerte y veloz.
- Linfocitos T supresores (o reguladores): Aunque son menos numerosos, estas células son importantes para "calmar" la respuesta inmunitaria. Ayudan a evitar que el sistema inmunitario reaccione de forma exagerada o ataque a las propias células del cuerpo, lo que podría causar enfermedades autoinmunes.
La producción de IL-2 por los linfocitos T colaboradores también es necesaria para que los linfocitos T CD8+ (los "soldados" que matan células infectadas) puedan multiplicarse. Sin la ayuda de los linfocitos T colaboradores, los linfocitos T CD8+ no podrían combatir eficazmente las infecciones.
Cómo los linfocitos T colaboradores dirigen la respuesta inmune
Los linfocitos T colaboradores son muy importantes porque pueden influir en cómo otras células inmunitarias responden a una amenaza. Para ser efectivos, deben decidir qué tipo de "mensajes" (citoquinas) enviar para que el sistema inmunitario sea lo más útil posible.
El modelo Th1/Th2
Los linfocitos T colaboradores se diferencian en dos tipos principales, conocidos como Th1 y Th2. Cada tipo se especializa en combatir diferentes tipos de amenazas:
| Tipo 1/ Th1 | Tipo 2/ Th2 | |
| Colabora principalmente con | Macrófagos (células que "comen" invasores). | Linfocito Bs (células que producen anticuerpos). |
| Citoquinas que produce | interferón gamma y factor de necrosis tumoral-beta. | interleucina-4, interleucina-5, interleucina-6, interleucina-10, interleucina-13. |
| Tipo de defensa que promueve | Inmunidad celular. Ayuda a los macrófagos a destruir invasores y a los linfocitos T citotóxicos a multiplicarse. Es buena contra virus y bacterias que viven dentro de las células. | Inmunidad humoral. Estimula a los linfocitos B para que produzcan muchos anticuerpos. Es buena contra bacterias y toxinas que están fuera de las células. |

Aunque este modelo Th1/Th2 nos ayuda a entender cómo funcionan los linfocitos T colaboradores, la realidad es más compleja. Hay otros tipos de células T y citoquinas que también influyen en la respuesta inmune, como los linfocitos T reguladores y los linfocitos Th17. Los científicos siguen investigando para comprender completamente cómo estas células deciden qué tipo de respuesta generar.
Papel de los linfocitos T en las enfermedades
Dado lo importantes que son los linfocitos T colaboradores, no es de extrañar que influyan mucho en cómo nuestro cuerpo combate las enfermedades. A veces, pueden cometer errores o generar respuestas que no son beneficiosas.
Linfocitos T colaboradores y reacciones de hipersensibilidad
El sistema inmunitario debe ser muy preciso: debe responder a los invasores, pero no atacar a las propias células del cuerpo. Cuando el sistema inmunitario reacciona de forma exagerada a sustancias que normalmente no son dañinas, se produce una reacción de hipersensibilidad. Esto es lo que ocurre en las alergias y algunas enfermedades autoinmunes.
Hay diferentes tipos de hipersensibilidad:
- Hipersensibilidad de tipo 1: Incluye alergias comunes como el asma, la rinitis alérgica (fiebre del heno) y las urticarias. En estas reacciones, los linfocitos Th2 son importantes para la producción de anticuerpos especiales (IgE) que causan los síntomas alérgicos.
- Hipersensibilidad de tipos 2 y 3: Involucran problemas con anticuerpos que atacan por error a las propias células o tejidos del cuerpo. Los linfocitos T colaboradores pueden influir en estas reacciones, por ejemplo, en enfermedades como el lupus.
- Hipersensibilidad de tipo 4: También conocida como hipersensibilidad tardía, es causada por una sobreestimulación de células inmunitarias como los linfocitos y los macrófagos, lo que lleva a inflamación crónica. Los linfocitos T son clave en este tipo, ya que se activan y promueven la acción de otras células, especialmente los macrófagos, a través de citoquinas Th1.
Los linfocitos T colaboradores también son necesarios para el desarrollo de algunas enfermedades autoinmunes donde los linfocitos T citotóxicos atacan por error a las propias células del cuerpo, como en la diabetes tipo 1. En estas enfermedades, el sistema inmunitario confunde una parte de nuestro cuerpo con un invasor y la ataca.
Infección por VIH
El VIH (virus de inmunodeficiencia humana) es un ejemplo muy claro de la importancia de los linfocitos T CD4+. El VIH ataca directamente a las células que tienen la proteína CD4, incluyendo los linfocitos T CD4+.
Al principio de la infección, el cuerpo intenta compensar la pérdida de estas células produciendo más. Sin embargo, con el tiempo, el virus se vuelve más agresivo y destruye los linfocitos T CD4+ más rápido de lo que el cuerpo puede producirlos.
Cuando el número de linfocitos T CD4+ funcionales disminuye drásticamente, el sistema inmunitario se debilita mucho, lo que lleva al sida. En esta etapa, el cuerpo no puede reconocer ni combatir eficazmente muchos patógenos, lo que permite que ocurran "infecciones oportunistas" (infecciones que normalmente no afectarían a una persona sana).
Dos partes del sistema inmunitario se ven muy afectadas por la falta de linfocitos T CD4+:
- Los linfocitos T CD8+ no se activan tan bien, lo que hace que las personas con sida sean más vulnerables a los virus, incluido el propio VIH. Esto permite que el virus se multiplique más rápido.
- La capacidad de los linfocitos B para producir anticuerpos de alta calidad disminuye. Esto hace que el cuerpo sea más susceptible a infecciones bacterianas.
Si no se recibe tratamiento para el VIH, el sistema inmunitario se debilita tanto que la persona puede sucumbir a infecciones o cánceres.
Galería de imágenes
-
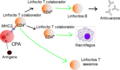
Función de los linfocitos (células) T: Las células presentadoras de antígeno (CPA) presentan el fragmento de antígeno sobre sus moléculas MHC de clase II (MHC II). Es entonces cuando los linfocitos T colaboradores reconocen ese complejo molecular, con la ayuda de la expresión de sus correceptores CD4 (CD4+). La activación de un linfocito T inactivo, ocasiona que libere citoquinas y otras señales de estimulación (flechas verdes) las cuales provocan un aumento en la actividad de macrófagos, linfocitos citotóxicos (CD8+) y linfocitos B, siendo estos últimos los que producen anticuerpos. Una imagen más detallada se encuentra abajo.
-
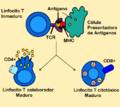
La presentación de antígenos estimula a los linfocitos (células) T a convertirse ya sean "citotóxicos" CD8+ o "colaboradores" CD4+.
Plantilla:Ficha de célula
Véase también
 En inglés: T helper cell Facts for Kids
En inglés: T helper cell Facts for Kids
