
Hemo para niños
El grupo hemo (que viene de la palabra griega "haima", que significa "sangre") es una parte muy importante de muchas proteínas. La más conocida es la hemoglobina, que es la que le da el color rojo a nuestra sangre y ayuda a transportar el oxígeno.
El grupo hemo está formado por un átomo de hierro (Fe) en el centro de una estructura orgánica grande llamada porfirina. Esta porfirina es como un anillo hecho de cuatro partes más pequeñas llamadas pirrol, unidas entre sí. No todas las porfirinas tienen hierro, pero muchas proteínas que sí lo tienen usan el grupo hemo. A estas proteínas se les llama hemoproteínas.
Además de la hemoglobina, el grupo hemo se encuentra en otras hemoproteínas importantes como la mioglobina (que guarda oxígeno en los músculos), los citocromos (que participan en la energía de las células) y la catalasa (que protege a las células).
Contenido
¿Para qué sirve el grupo hemo?

Las hemoproteínas tienen varias funciones vitales en nuestro cuerpo:
- Transporte de gases: Llevan gases como el oxígeno.
- Ayuda en reacciones químicas: Actúan como catalizadores, acelerando procesos químicos.
- Detección de gases: Pueden sentir la presencia de ciertos gases.
- Transferencia de electrones: El hierro en el grupo hemo puede ganar o perder electrones, lo cual es clave en muchas reacciones de energía.
Cuando el grupo hemo detecta un gas, como el oxígeno, el gas se une al hierro. Esto hace que la proteína cambie de forma, lo que le permite cumplir su función.
Se cree que, hace mucho tiempo, la función original de las hemoproteínas era ayudar a mover electrones en la fotosíntesis de organismos muy antiguos, antes de que hubiera mucho oxígeno en la Tierra.
Las hemoproteínas han evolucionado para tener muchas funciones diferentes. Por ejemplo, la hemoglobina puede entregar oxígeno a los tejidos de nuestro cuerpo de manera muy eficiente. Esto se debe a que tiene unas partes especiales llamadas aminoácidos cerca del grupo hemo. La hemoglobina toma oxígeno en los pulmones y lo suelta en los tejidos, donde se necesita. Este proceso se conoce como el efecto Bohr.
¿Cómo es la estructura química del grupo hemo?
El grupo hemo está formado por un átomo de hierro (Fe) en el centro de un anillo de porfirina. Este anillo es un tetrapirrol cíclico, lo que significa que está compuesto por cuatro anillos de pirrol unidos entre sí. El átomo de hierro se une a cuatro nitrógenos que están en el centro del anillo de porfirina.
Tipos de grupos hemo
Existen varios tipos de grupos hemo que son importantes en la biología. Los más comunes son el Hemo B, el Hemo A y el Hemo C.
| Hemo A | Hemo B | Hemo C | Hemo O | ||
|---|---|---|---|---|---|
| Fórmula química | C 49H 56O 6N 4Fe |
C 34H 32O 4N 4Fe |
C 34H 36O 4N 4S 2Fe |
C 49H 58O 5N 4Fe |
|
| Grupo funcional en C 3 |
 |
-CH(OH)-CH 2-Far |
-CH=CH 2 |
-CH(cisteín-S-il)-CH 3 |
-CH(OH)-CH 2-Far |
| Grupo funcional en C 8 |
-CH=CH 2 |
-CH=CH 2 |
-CH(cisteín-S-il)-CH 3 |
-CH=CH 2 |
|
| Grupo funcional en C 18 |
-CH=O | -CH 3 |
-CH 3 |
-CH 3 |
|


El Hemo B es el tipo más común en la naturaleza. Los nombres de los citocromos suelen indicar qué tipo de grupo hemo contienen. Por ejemplo, el citocromo a tiene Hemo A, y el citocromo c tiene Hemo C.
Otros tipos de hemo
Existen otros tipos de hemo menos comunes pero también importantes:
- Hemo l: Se encuentra en algunas enzimas protectoras de animales, como la lactoperoxidasa, que ayudan a defender el cuerpo contra organismos invasores.
- Hemo m: Está en la mieloperoxidasa, una enzima que se encuentra en nuestras células de defensa (neutrófilos) y ayuda a destruir bacterias y virus.
- Hemo D: Es importante en algunas bacterias para ayudar en la respiración cuando hay poco oxígeno.
- Hemo S: Se encuentra en la hemoglobina de algunos gusanos marinos.
¿Cómo se forma el grupo hemo?

El proceso para producir el grupo hemo se llama porfirinosíntesis. Este proceso es muy similar en todos los seres vivos. En los humanos, casi toda la porfirinosíntesis se usa para crear hemo.
La producción de hemo comienza con la formación de una molécula llamada ácido δ-aminolevulínico (dALA). Una enzima clave en este paso, la ALA sintasa, es controlada por la cantidad de hemo que ya hay en el cuerpo. Si hay mucho hemo, la producción se ralentiza.
Aunque todas nuestras células necesitan hemo, los órganos principales que lo producen son el hígado y la médula ósea. El hígado ajusta su producción según las necesidades del cuerpo, mientras que la médula ósea lo produce de forma más constante para la hemoglobina.
Si hay problemas con las enzimas que participan en la síntesis del hemo, pueden aparecer enfermedades llamadas porfirias.
¿Cómo se descompone el grupo hemo?

Cuando los glóbulos rojos envejecen, son retirados de la circulación por unas células especiales en el bazo. Allí, el grupo hemo se descompone.
1. Primero, el hemo se convierte en biliverdina gracias a una enzima llamada hemo oxigenasa. En este paso, se libera monóxido de carbono (CO) y el hierro del hemo. El monóxido de carbono puede actuar como una señal en el cuerpo. 2. Luego, la biliverdina se transforma en bilirrubina por otra enzima. 3. La bilirrubina viaja al hígado, donde se modifica para que sea más fácil de eliminar. 4. Finalmente, esta bilirrubina modificada se expulsa del hígado a través de la bilis. Una parte se elimina por la orina (dándole su color amarillo) y otra parte por las heces (dándoles su color oscuro).
La descomposición del hemo también es una forma en que las células se protegen del estrés. Si hay muchos radicales libres (sustancias que pueden dañar las células), la célula aumenta su capacidad para descomponer el hemo, lo que ayuda a limitar los efectos dañinos del hemo libre.
El grupo hemo en la salud
Normalmente, el grupo hemo está bien protegido dentro de las proteínas. Pero si hay mucho estrés en el cuerpo, algunas proteínas pueden liberar sus grupos hemo. El hemo "libre" (que no está unido a una proteína) puede ser dañino porque el hierro que contiene puede producir radicales libres, que son moléculas que pueden dañar las células. Esto puede jugar un papel en algunas enfermedades.
Genes relacionados con el hemo
Algunos genes importantes que participan en la formación del hemo son:
- ALAD
- ALAS1
- ALAS2
- CPOX
- FECH
- HMBS
- PPOX
- UROD
- UROS
Galería de imágenes
-
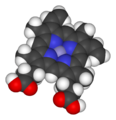
Grupo Hemo B. Se aprecia el ion ferroso (Fe2+
) en el centro de la protoporfirina IX. -
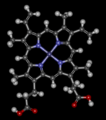
Modelo de Bolas y Bastones del Hemo B
-
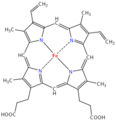
Grupo Hemo de la hemoglobina. Un átomo de hierro (Fe) en el centro aparece en rojo, formando complejo con cuatro átomos de nitrógeno interiores que aparecen en azul. Debajo, los dos ácidos carboxílicos COOH.



Véase también
 En inglés: Haem Facts for Kids
En inglés: Haem Facts for Kids
