
Homología (biología) para niños
En el estudio de los seres vivos, la homología es la relación que existe entre dos partes del cuerpo de diferentes organismos cuando estas partes tienen el mismo origen evolutivo.
Esto significa que dos órganos de dos especies distintas son homólogos si ambos provienen de un órgano similar en su antepasado común, sin importar cuánto hayan cambiado con el tiempo. Por ejemplo, las cuatro extremidades de los vertebrados con mandíbula, desde los tiburones hasta las aves o los mamíferos, son homólogas. De la misma manera, el extremo de la pata de un caballo es homólogo al dedo medio de la mano y el pie humano.
En la evolución, los rasgos homólogos se pueden rastrear hasta un mismo rasgo en el antepasado común de los grupos de seres vivos que los poseen.
Contenido
Homología, analogía y homodinamia
¿Qué diferencia hay entre homología y analogía?
Una homología muestra que dos estructuras tienen la misma combinación genética y provienen de un antepasado común.
Por otro lado, una analogía es una estructura que se parece a otra o que cumple la misma función, pero su desarrollo embrionario y su origen son diferentes. Las analogías no se encuentran en un antepasado común; son el resultado de la convergencia evolutiva, donde diferentes especies desarrollan características similares de forma independiente para adaptarse a entornos parecidos.
| Estructuras | Procesos | |
|---|---|---|
 |
Homología Dos estructuras son homólogas si son parecidas en su forma y si esta semejanza se debe a que vienen de una estructura ancestral común. Un ejemplo es la comparación de las alas del pterodactylus y del murciélago. |
Paralelismo. El paralelismo ocurre cuando procesos de desarrollo similares llevan a estructuras parecidas. La biología evolutiva del desarrollo estudia esto. |
 |
Analogía Dos estructuras son análogas si son parecidas en su forma o función, y si esta semejanza se ha logrado de forma independiente en la evolución. Por ejemplo, las alas de las mariposas y las alas de los murciélagos y las aves. |
Convergencia La convergencia evolutiva es el resultado de presiones selectivas similares que hacen que especies diferentes desarrollen rasgos parecidos. |
¿Qué es la homodinamia?
La homodinamia se refiere a cuando ciertas señales en el desarrollo de un embrión tienen las mismas consecuencias en dos organismos diferentes. Por ejemplo, si la piel embrionaria de una rana se trasplanta a un embrión de salamandra en la zona donde se formaría la mandíbula, y produce una mandíbula de rana en la cabeza de la salamandra, eso es homodinamia. Significa que ambos organismos tienen una capacidad similar para responder de la misma manera a una señal de desarrollo.
Ejemplos de homología en animales
Uno de los ejemplos más claros de homología es la relación entre el cartílago branquial de los peces, la mandíbula de los reptiles y los huesos del oído medio de los mamíferos:
- Los arcos branquiales de los peces sin mandíbula evolucionaron para formar la mandíbula de los peces con mandíbula. Estos arcos estaban debajo de la boca y, al sujetarse con cartílagos, rodearon la boca para formar la mandíbula.
- La parte superior del segundo arco branquial, que sostenía la branquia, se convirtió en el hueso hiomandibular. Este hueso sostenía el cráneo, uniéndolo a la mandíbula. Cuando los vertebrados se adaptaron a la tierra, enfrentaron nuevos desafíos, como escuchar en el aire. El hueso hiomandibular era ideal para esta función, ya que estaba cerca del tímpano y los huesos son buenos para transmitir el sonido. Así, este hueso empezó a tener una doble función: seguir sosteniendo el cráneo y transmitir el sonido. A medida que los vertebrados terrestres cambiaron su postura y estructura de la mandíbula, el cráneo ya no necesitó el soporte del hueso hiomandibular, que pudo especializarse y convertirse en el estribo (un hueso del oído).
- En anfibios, reptiles y aves, la parte trasera del cartílago forma el hueso cuadrático de la mandíbula superior y el hueso articular de la mandíbula inferior. Estos huesos se conectan y permiten el movimiento de la mandíbula. Sin embargo, en los mamíferos, esta articulación ocurre en otra parte, lo que liberó a estos huesos para adquirir nuevas funciones. El hueso cuadrático de la mandíbula superior de los reptiles evolucionó en el hueso incus (yunque) del oído medio de los mamíferos, y el hueso articular de la mandíbula inferior de los reptiles se convirtió en el malleus (martillo).
Ejemplos de rasgos homólogos
- Los brazos humanos y las aletas de las ballenas tienen el mismo antepasado común, aunque no cumplen la misma función.
Ejemplos de rasgos análogos
- Las alas de las aves y las de los insectos.
- La aleta caudal (cola) de los peces y la de las ballenas.
- Las patas de los artrópodos (como los insectos) y las de los vertebrados.
Homología especial y homología serial

También podemos hablar de homología dentro de un mismo organismo o especie. Existe homología serial entre órganos que se repiten, como las diferentes hojas de una planta o los tres pares de patas de un insecto. En este sentido, el pulgar de la mano y el dedo gordo del pie son homólogos.
El concepto de homología a lo largo del tiempo
El concepto más común de homología es el histórico: dos órganos son homólogos cuando provienen de un "órgano ancestral" común. Esta definición fue propuesta por Ray Lankester para aclarar la idea de Richard Owen, quien definía la homología como "el mismo órgano en diferentes animales". Lankester especificó que los rasgos homólogos son aquellos que "tienen un solo representante en un antepasado común".
La biología evolutiva del desarrollo usa una definición diferente de homología que considera los procesos de desarrollo que dan origen a los órganos homólogos. Este nuevo concepto se llama homología ontogenética o concepto biológico de homología. Desde esta perspectiva, la homología no es solo una característica pasiva, sino que puede influir en la trayectoria evolutiva.
Cómo se comprueba la homología
Como las afirmaciones de homología son hipótesis, se han propuesto tres pruebas para verificarlas:
Similitud
Es la prueba más antigua y se basa en tres criterios: la posición o relación en el cuerpo, la similitud estructural y la conexión entre formas intermedias. Los dos primeros se usan más a menudo. Algunos científicos creen que la similitud por sí sola no es una prueba fuerte de homología, ya que estructuras no homólogas también pueden ser similares.
Conjunción
Esta prueba descarta como homólogos a dos rasgos similares que se encuentran juntos en los mismos organismos.
Congruencia
Es la prueba más importante de la homología. Si una estructura que se cree homóloga no encaja con otras estructuras homólogas en el árbol evolutivo, se considera que no pasó la prueba de congruencia.
Genética y homología
A partir de los años 20, la homología empezó a verse desde una perspectiva genética. En 1920, Alexander Weinstein usó el término genes homólogos para referirse a genes de especies diferentes con características fenotípicas similares. En 1934, Alan Boyden propuso usar la genética para reconocer homologías, considerándolas por primera vez como un "fenómeno genético". Al principio, se consideraba que dos genes eran homólogos solo si tenían la misma característica visible. Sin embargo, a partir de los años sesenta, se adoptó la idea actual de homología genética: dos genes de dos especies distintas son homólogos si provienen de un mismo gen ancestral.
Homología de secuencia

La secuencia de nucleótidos de un gen se transmite de padres a hijos y es lo que cambia principalmente con la evolución. Cuando estudiamos el genoma de dos especies, esperamos encontrar los genes equivalentes en ambas, con una secuencia algo diferente. Cuanto más lejano sea el antepasado común, más diferentes serán las secuencias. La expresión homología de secuencia se refiere a la correspondencia entre las cadenas de nucleótidos de esos dos genes, lo que nos permite reconocer que son homólogos.
Genes ortólogos y genes parálogos
Dentro de la homología de secuencia, hay dos tipos:
- Los genes ortólogos son similares porque pertenecen a dos especies que tienen un antepasado común.
- Los genes parálogos se encuentran en el mismo organismo, y su similitud indica que uno se originó por la duplicación del otro.
La duplicación génica es un evento evolutivo importante. Una vez que un gen se duplica, las copias evolucionan por separado y pueden dar lugar a productos diferentes, abriendo camino a nuevas adaptaciones. En biología molecular, la paralogía es similar a la homología serial. Por ejemplo, los genes que determinan los diferentes tipos de hemoglobinas que se producen a lo largo de la vida fetal y adulta son parálogos. La hemoglobina tiene cuatro globinas. En los vertebrados primitivos, estas cuatro cadenas eran del mismo tipo. Sin embargo, en los vertebrados superiores, la hemoglobina tiene dos cadenas de globina α y β, debido a una duplicación genética que llevó a dos copias del gen de globina original. Ambas copias cambiaron a lo largo de la evolución, dando lugar a dos genes de globina especializados y sus productos.
GenBank es una base de datos donde se guardan todas las secuencias de ADN. Para buscar homologías, se usa una herramienta llamada BLAST. Se introduce una secuencia y se obtiene una lista de todas las secuencias guardadas que se parecen a la introducida, ordenadas por grado de similitud.
Homología estructural y homología de procesos
Entre las homologías moleculares descubiertas, las más sorprendentes fueron las relacionadas con genes reguladores como los genes Hox. Edward B. Lewis y su equipo fueron los primeros en analizar una región del cromosoma 3 de la mosca Drosophila melanogaster que contenía varios genes homeóticos, que más tarde se encontraron también en los vertebrados.
El descubrimiento de estas homologías moleculares en la biología evolutiva del desarrollo ha hecho que se reconsidere la diferencia clásica entre homología y analogía. Un ejemplo son los ojos y los corazones de vertebrados e insectos, que clásicamente se consideraban ejemplos de analogía:
- Recientemente, se ha demostrado que la formación de los ojos de insectos y vertebrados depende de la acción de genes homólogos (Pax-6 en el ratón y eyeless en la Drosophila). Esto sugiere que los ojos de vertebrados, insectos y cefalópodos podrían descender de una célula básica fotorreceptiva regulada por el mismo gen.
- Desde el punto de vista de su forma, los corazones de vertebrados e insectos solo tienen en común que bombean líquidos, por lo que se consideraban un ejemplo de analogía. Sin embargo, se ha demostrado que ambos dependen de la acción de un mismo gen (Csx/tinman).
Aun así, muchos biólogos han sido cautelosos con el entusiasmo por las homologías moleculares y no han querido redefinir la homología solo en términos genéticos. El argumento principal es que los genes homólogos pueden activarse en estructuras que no son homólogas. Este es el caso de los genes Hox: su descubrimiento en ratones y moscas llevó a pensar que el control de la segmentación del cuerpo era homólogo en artrópodos y vertebrados. Sin embargo, se ha visto que los complejos Hox también pueden organizar otras partes del cuerpo, como las extremidades. Algunos científicos han propuesto que la homología es un fenómeno que ocurre en diferentes niveles de organización del cuerpo. Así, el ojo de vertebrados y artrópodos serían homólogos como órganos que detectan la luz, pero no como estructuras complejas y organizadas. En la misma línea, Scott Gilbert ha propuesto el concepto de homología de procesos para los patrones de expresión genética homólogos, y mantener el concepto de homología estructural para las homologías clásicas.
La homología de las alas de las aves ha sido muy importante para los paleontólogos y biólogos evolutivos, ya que está ligada a la evolución de las aves. Es un ejemplo clásico de desacuerdos entre la anatomía y la embriología. En la biología del desarrollo, los dedos de las alas de las aves se consideran, basándose en el desarrollo embrionario, los dedos 2, 3 y 4. En contraste, la paleontología, basándose en el análisis de fósiles, sugiere que los dedos de las alas son los 1, 2 y 3, indicando que las aves descienden de dinosaurios terópodos que perdieron los dedos 4 y 5. Se ha argumentado que el desarrollo de las alas no apoya la idea de que las aves son terópodos, y que las aves descienden de un antepasado que perdió los dedos 1 y 5. Sin embargo, estudios en el desarrollo de las extremidades de ratones y pollos han concluido que el dedo 1 en desarrollo no expresa el gen Hoxd12, pero sí el Hoxd13. El resto de los dedos expresan tanto Hoxd12 como Hoxd13. Este patrón de expresión identifica el dedo más delantero del ala como el dedo 1. De acuerdo con la hipótesis anterior, estos dedos son el 1, 2 y 3, como en los dinosaurios terópodos, lo que apoya que las aves son dinosaurios. Gracias a toda la evidencia experimental, se llegó al acuerdo de que hubo un cambio en la identidad de los dedos, es decir, una homeosis (cambio en la identidad de una parte del cuerpo) en la evolución de los dedos de las aves actuales.
Un caso similar al de las aves es el del lagarto Chalcides chalcides, que tiene extremidades reducidas con tres dedos en lugar de cinco, como la mayoría de los lagartos de su grupo. Estudios de anatomía indican que los tres dedos de las extremidades delanteras de esta especie corresponden a los dedos 1, 2 y 3, mientras que estudios de desarrollo sugieren que corresponden a los 2, 3 y 4. Mediante la expresión del gen Hoxd11, se estableció que los dedos 1, 2 y 3 se desarrollan en las posiciones 2, 3 y 4. Esto sugiere que, al igual que en las aves, en estos lagartos también ocurrió un patrón de cambio homeótico.
Ejemplo: el origen de las extremidades de los animales
El gen distal-less produce una proteína que es clave en la organización del crecimiento y la formación de los patrones de las patas de la mosca Drosophila melanogaster. La acción de este mismo gen también es fundamental en el desarrollo de las extremidades y aletas de los cordados (animales con columna vertebral), en las patas de los poliquetos (gusanos marinos), en los lobópodos de los onicóforos (gusanos aterciopelados), en las ampollas de las ascidias (animales marinos) y en el aparato ambulacral de los equinodermos (como las estrellas de mar). La pregunta evolutiva es si podemos considerar que estas extremidades son homólogas y que, por lo tanto, provienen de una extremidad de un antepasado común a estos seis grupos. Sin embargo, el registro fósil muestra claramente que el antepasado común de todos estos grupos no tenía extremidades. Por lo tanto, las extremidades de artrópodos, anélidos, equinodermos y cordados, como estructuras físicas (homología estructural), no son homólogas. Pero la secuencia y la acción del gen distal-less, así como su función en la formación de las extremidades, son homólogas en todos estos grupos (homología de procesos).
El método comparativo
El concepto de homología es central en el método comparativo que se usa en biología (puedes ver el artículo Anatomía comparada). Desde que Darwin explicó cómo se originó la diversidad de la vida, la comparación se hace principalmente entre partes homólogas de los organismos. La comparación homóloga es la única válida en el análisis filogenético (llamado cladística), que busca entender la historia evolutiva de las formas de vida.
Galería de imágenes
-

Homología de varios huesos (mostrados en distintos colores) de las extremidades delanteras de cuatro vertebrados.
-
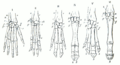
Carl Gegenbaur: Homología entre miembros anteriores (1870).


Véase también
 En inglés: Homology Facts for Kids
En inglés: Homology Facts for Kids
