
Endosimbionte para niños
Un endosimbionte es un organismo que vive dentro del cuerpo o las células de otro organismo, llamado huésped. A menudo, esta relación es de mutualismo, lo que significa que ambos organismos se benefician. La palabra "endosimbiosis" viene del griego: endon (dentro), syn (juntos) y biosis (vivo).
Algunos ejemplos de endosimbiontes son:
- Bacterias que fijan nitrógeno (llamadas rizobios) que viven en las raíces de plantas como los guisantes.
- Algas unicelulares que viven dentro de los corales que forman arrecifes.
- Bacterias que proporcionan nutrientes esenciales a muchos insectos.
Contenido
- ¿Cómo se transmiten los endosimbiontes?
- Endosimbiosis obligatoria
- Simbiogénesis y orgánulos celulares
- Endosimbiontes en invertebrados
- Endosimbiontes del fitoplancton
- Endosimbiontes de protistas
- Endosimbiontes de vertebrados
- Endosimbiontes de plantas
- Endosimbiontes de bacterias
- Asociaciones virus-huésped
- Galería de imágenes
- Véase también
¿Cómo se transmiten los endosimbiontes?
Existen dos formas principales en que los endosimbiontes pasan de un organismo a otro:
Transmisión horizontal
En este tipo, cada nueva generación de un organismo adquiere sus simbiontes del ambiente. Por ejemplo, las bacterias que fijan nitrógeno en las raíces de algunas plantas se obtienen del suelo.
Transmisión vertical
Aquí, el simbionte se transfiere directamente de los padres a sus hijos. Cuando esto ocurre, los simbiontes a menudo pierden algunos de sus genes y ya no pueden vivir por sí mismos. Esto crea una relación de gran dependencia, donde el simbionte necesita al huésped para sobrevivir, y el huésped a menudo necesita al simbionte para obtener nutrientes importantes.
Por ejemplo, las bacterias que viven dentro de los pulgones (pequeños insectos que comen plantas) han perdido genes esenciales. Ahora dependen del pulgón para obtener ciertos nutrientes. A cambio, estas bacterias producen aminoácidos que el pulgón necesita para vivir.
Cuando un simbionte se vuelve tan dependiente y vive tan íntimamente dentro de las células de su huésped, puede empezar a parecerse a un orgánulo celular, como las mitocondrias o los cloroplastos.
Endosimbiosis obligatoria
Muchos casos de endosimbiosis son "obligatorios". Esto significa que el endosimbionte o el huésped no pueden sobrevivir el uno sin el otro. Un ejemplo son los gusanos marinos del género Riftia, que no tienen boca ni sistema digestivo y obtienen todos sus nutrientes de las bacterias que viven dentro de ellos.
Los ejemplos más conocidos de endosimbiosis obligatoria son las mitocondrias y los cloroplastos, que son partes esenciales de nuestras células y de las células de las plantas.
Algunos microorganismos que afectan a otros seres vivos, como Wuchereria bancrofti y Mansonella perstans, dependen de una bacteria llamada Wolbachia spp. para vivir en sus huéspedes insectos. Si se elimina esta bacteria, los microorganismos no pueden sobrevivir.
Simbiogénesis y orgánulos celulares

La simbiogénesis es la teoría que explica cómo surgieron las células eucariotas (las células que tienen un núcleo definido, como las de los animales, plantas y hongos). Esta teoría propone que las mitocondrias y los cloroplastos, dos tipos importantes de orgánulos dentro de las células eucariotas, evolucionaron a partir de bacterias.
Se cree que hace mucho tiempo, una célula eucariota primitiva "engulló" a ciertas bacterias. En lugar de digerirlas, estas bacterias se quedaron dentro de la célula y establecieron una relación endosimbiótica. Con el tiempo, estas bacterias se adaptaron a vivir exclusivamente dentro de las células eucariotas, convirtiéndose en las mitocondrias (que producen energía) y los cloroplastos (que realizan la fotosíntesis en las plantas).
Muchos insectos tienen endosimbiontes que están en diferentes etapas de esta simbiogénesis. A menudo, los genes de los simbiontes se reducen a solo los más esenciales para la vida en conjunto con el huésped.
Endosimbiontes en invertebrados
Los casos de endosimbiosis mejor estudiados se encuentran en los invertebrados (animales sin columna vertebral). Estas relaciones son muy importantes para organismos como los corales (con sus algas Symbiodinium) o los insectos (con bacterias como Wolbachia).
Endosimbiontes de insectos
_-_with_key.png)
Los endosimbiontes de insectos se dividen en dos categorías:
Endosimbiontes primarios
Estos simbiontes han vivido con sus insectos huéspedes durante millones de años. Su relación es obligatoria, lo que significa que ni la bacteria ni el insecto pueden vivir el uno sin el otro. Además, han evolucionado juntos, un proceso llamado coespeciación.
Un ejemplo famoso es el pulgón del guisante (Acyrthosiphon pisum) y su bacteria Buchnera. La Buchnera ayuda al pulgón a producir aminoácidos esenciales que no puede obtener de su dieta de savia de plantas. Otro ejemplo es la mosca tsetsé y su bacteria Wigglesworthia, que le proporciona vitaminas.
Estas bacterias suelen vivir en células especiales del insecto llamadas bacteriocitos y se transmiten de la madre a sus crías. Las bacterias se benefician de un ambiente seguro, con muchos nutrientes y poca competencia.
Los científicos han descubierto que los genomas (el conjunto de genes) de estas bacterias obligadas son muy pequeños. Han perdido muchos genes que otras bacterias tienen, porque ya no los necesitan dentro de la célula del insecto. Estudiar estos simbiontes es importante para entender cómo controlar plagas de insectos o insectos que transmiten enfermedades, ya que al atacar a la bacteria, se puede afectar al insecto.
Endosimbiontes secundarios
Estos simbiontes tienen una relación más reciente con sus huéspedes y a veces pueden transferirse entre diferentes insectos. No siempre son obligatorios.
Por ejemplo, el pulgón del guisante también puede tener otros endosimbiontes secundarios como Hamiltonella defensa. Esta bacteria ayuda al pulgón a defenderse de avispas que intentan poner sus huevos dentro de él.
Otro simbionte defensivo es Spiroplasma poulsonii, que protege a las moscas Drosophila de parásitos. Produce toxinas que atacan a los parásitos invasores.
Endosimbiontes de hormigas
Las hormigas del género Camponotus tienen una bacteria llamada Blochmannia como su endosimbionte principal. En 2018, se descubrió otro simbionte primario en las hormigas Cardiocondyla, llamado Candidatus Westeberhardia Cardiocondylae.
Endosimbiontes de invertebrados marinos
Muchos animales marinos como los equinodermos (estrellas de mar, erizos de mar) y algunos gusanos marinos tienen endosimbiontes. Algunos gusanos marinos, como Olavius algarvensis, dependen completamente de bacterias que producen su propio alimento (quimioautotróficas) y no tienen sistema digestivo.
Endosimbiontes de dinoflagelados
Los dinoflagelados del género Symbiodinium, también conocidos como zooxantelas, son endosimbiontes muy importantes para los corales, moluscos (como las almejas gigantes) y esponjas. Estos simbiontes realizan la fotosíntesis (usan la luz solar para crear energía) y se la proporcionan a sus huéspedes, lo que es crucial para la formación de los arrecifes de coral.
Endosimbiontes del fitoplancton
En el océano, se han descubierto endosimbiontes bacterianos en el fitoplancton (pequeños organismos que flotan en el agua y hacen fotosíntesis). Estas relaciones son comunes en zonas del océano con pocos nutrientes. Las bacterias endosimbióticas fijan nitrógeno para sus huéspedes, como las diatomeas, y a cambio reciben carbono orgánico de la fotosíntesis. Esto es muy importante para el ciclo global del carbono en estas áreas.
Un ejemplo es la relación entre la diatomea Hemialus y la cianobacteria Richelia intracellularis. La Richelia vive dentro de la diatomea y le proporciona nitrógeno, lo que permite que ambos crezcan mucho más que si vivieran solos.
Endosimbiontes de protistas
Los protistas son organismos unicelulares muy diversos. Algunos tienen endosimbiontes:
- Mixotricha paradoxa es un protozoo que no tiene mitocondrias, pero tiene bacterias esféricas dentro de su célula que cumplen esa función.
- Paramecium bursaria, un tipo de ciliado, vive en una relación mutualista con un alga verde llamada Zoochlorella que vive dentro de su citoplasma.
- Paulinella chromatophora es un ameboide de agua dulce que recientemente adquirió una cianobacteria como endosimbionte.
- Muchos foraminíferos (pequeños organismos marinos con caparazones) albergan diferentes tipos de algas.
- Hatena arenicola es un protista que, cuando engulle un alga verde, deja de alimentarse y se vuelve fotosintético.
Endosimbiontes de vertebrados
La salamandra manchada (Ambystoma maculatum) tiene una relación con el alga Oophila amblystomatis, que crece en las cápsulas de sus huevos.
Endosimbiontes de plantas
Los cloroplastos son los endosimbiontes primarios de las plantas. Producen energía para la planta mediante la fotosíntesis, creando azúcares.
La planta acuática Azolla tiene una relación muy cercana con su simbionte, una cianobacteria llamada Anabaena, que se transmite directamente de una generación a la siguiente.
Endosimbiontes de bacterias
Incluso algunas bacterias pueden tener endosimbiontes. Se ha observado que algunas Betaproteobacterias tienen Gammaproteobacterias viviendo dentro de ellas.
Asociaciones virus-huésped
El estudio del genoma humano ha revelado miles de retrovirus endógenos. Estos son elementos virales que se han integrado en nuestro genoma a lo largo de la evolución y se parecen mucho a los retrovirus.
Galería de imágenes
-
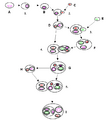
Una representación de la teoría endosimbiótica
-

Los pulgones de los guisantes suelen estar infestados por avispas parásitas. Sus endosimbiontes secundarios atacan a las larvas de avispa parasitoide infestantes promoviendo la supervivencia tanto del pulgón huésped como de sus endosimbiontes.


Véase también
 En inglés: Endosymbiosis Facts for Kids
En inglés: Endosymbiosis Facts for Kids
