
Transcripción genética para niños
La transcripción del ADN es un proceso fundamental en nuestras células. Imagina que el ADN es un gran libro de recetas que contiene todas las instrucciones para construir y hacer funcionar nuestro cuerpo. La transcripción es el primer paso para usar esas recetas.
En este proceso, la información guardada en una parte del ADN (un gen) se copia a una molécula diferente llamada ARN. Piensa en el ARN como una copia de trabajo o un mensaje corto de una receta específica. Esta copia de ARN luego se usa para crear las proteínas, que son como los "ladrillos" y "herramientas" que construyen y realizan las funciones en nuestro cuerpo.
Una enzima muy importante llamada ARN polimerasa es la encargada de hacer esta copia. La ARN polimerasa lee la secuencia del ADN y construye una nueva cadena de ARN que lleva la misma información.
Contenido
Pasos de la Transcripción
La transcripción sigue varios pasos clave:
- La ARN polimerasa y otras proteínas especiales se unen a una parte del ADN llamada promotor, que es como la señal de "inicio" de un gen.
- La ARN polimerasa abre la doble hélice del ADN, separando sus dos cadenas. Esto crea una pequeña "burbuja" donde el ADN está abierto.
- La ARN polimerasa empieza a añadir "letras" de ARN (llamadas nucleótidos) que son complementarias a una de las cadenas de ADN.
- Estas "letras" de ARN se unen para formar una nueva cadena de ARN.
- La cadena de ARN recién hecha se separa del ADN.
- Si la célula tiene un núcleo (como las nuestras), el ARN puede ser modificado antes de salir.
- Finalmente, el ARN puede quedarse en el núcleo o ir al citoplasma para seguir con el siguiente paso: la fabricación de proteínas.
Etapas Principales
La transcripción se divide en tres etapas principales:
Configuración y Preparación
Antes de que la transcripción comience, varias proteínas llamadas factores de transcripción se unen a secuencias específicas en el ADN, especialmente en una zona llamada promotor. El promotor es como el punto de partida para la ARN polimerasa. Estas proteínas ayudan a la ARN polimerasa a encontrar el lugar correcto para empezar a copiar el gen.
Iniciación: El Comienzo de la Copia
Una vez que la ARN polimerasa y los factores de transcripción están en su lugar, la ARN polimerasa abre la doble hélice del ADN en la región del promotor. Esta apertura se llama "burbuja de transcripción". La ARN polimerasa entonces empieza a añadir las primeras "letras" de ARN, formando una pequeña cadena. Cuando se forma el primer enlace entre estas "letras", la etapa de iniciación termina.
Disgregación del Promotor
Después de que la ARN polimerasa ha comenzado a copiar y la cadena de ARN tiene una cierta longitud, el complejo que estaba unido al promotor se deshace. Esto permite que el promotor quede libre para que otra ARN polimerasa pueda iniciar una nueva transcripción.
Elongación: La Copia Continúa
En esta etapa, la ARN polimerasa se mueve a lo largo de una de las cadenas de ADN (la cadena molde) y sigue añadiendo "letras" de ARN una por una. La ARN polimerasa se asegura de que cada "letra" de ARN sea la correcta, complementaria a la "letra" del ADN. A medida que la ARN polimerasa avanza, la cadena de ARN se va haciendo más larga. Es importante saber que, aunque el ADN tiene timina (T), el ARN tiene uracilo (U) en su lugar.
Terminación: El Final de la Copia
Cuando la ARN polimerasa llega al final del gen, encuentra una señal especial en el ADN que le indica que debe detenerse. Esta señal hace que la ARN polimerasa se separe del ADN y que la cadena de ARN recién sintetizada se libere. El ADN vuelve a cerrarse y la ARN polimerasa queda libre para empezar otra transcripción.
Transcripción en Bacterias (Procariotas)
Las bacterias son organismos muy pequeños que no tienen núcleo. En ellas, la transcripción ocurre en el citoplasma.
La ARN Polimerasa Bacteriana
La ARN polimerasa en bacterias tiene varias partes. Una parte importante es el "factor sigma", que ayuda a la enzima a encontrar el lugar exacto en el ADN donde debe empezar a copiar. Una vez que la copia ha comenzado, el factor sigma se separa y la ARN polimerasa continúa sola. Esta enzima también tiene una función de "revisión" para corregir algunos errores.
Inicio de la Copia en Bacterias
El factor sigma de la ARN polimerasa se une a unas secuencias especiales en el ADN llamadas regiones promotoras. Estas regiones son como las "instrucciones de inicio" para la enzima. Una vez unida, la ARN polimerasa abre el ADN y comienza a copiar. A veces, produce cadenas de ARN muy cortas que no son útiles, pero finalmente logra hacer una cadena más larga y funcional.
Alargamiento de la Cadena en Bacterias
La ARN polimerasa se desliza por el ADN, desenrollándolo y copiando su información en una nueva cadena de ARN. Los nuevos nucleótidos de ARN se añaden al final de la cadena que está creciendo. Este proceso es muy rápido, y varias ARN polimerasas pueden estar copiando el mismo gen al mismo tiempo, lo que permite producir muchas moléculas de ARN rápidamente.
Final de la Copia en Bacterias
La transcripción en bacterias puede terminar de dos maneras:
- Terminación intrínseca: Algunas secuencias en el ADN hacen que el ARN recién formado se pliegue en una forma de "horquilla". Esta forma desestabiliza la unión entre el ARN y el ADN, haciendo que la ARN polimerasa se detenga y se libere.
- Terminación dependiente de Rho: Una proteína llamada factor Rho se une a la cadena de ARN y sigue a la ARN polimerasa. Cuando la polimerasa se detiene en ciertas secuencias, el factor Rho la alcanza y ayuda a separar el ARN del ADN, terminando la transcripción.
Transcripción en Células Eucariotas
Las células eucariotas (como las de los animales, plantas y hongos) son más complejas y tienen un núcleo. En ellas, la transcripción ocurre dentro del núcleo.
Tipos de ARN Polimerasa en Eucariotas
A diferencia de las bacterias, las células eucariotas tienen tres tipos principales de ARN polimerasa, cada una con una función específica:
- ARN Polimerasa I (Pol I): Copia los genes que forman la mayor parte del ARN ribosomal, que es un componente de los ribosomas (las "fábricas" de proteínas de la célula).
- ARN Polimerasa II (Pol II): Es la más importante para nosotros, ya que copia los genes que se convertirán en ARNm. El ARNm lleva las instrucciones para construir proteínas. También copia otros tipos de ARN pequeños.
- ARN Polimerasa III (Pol III): Copia los genes que forman el ARNt (que ayuda a llevar los aminoácidos para construir proteínas) y otros ARN pequeños.
Inicio de la Copia en Eucariotas
El inicio de la transcripción en eucariotas es más complejo. La ARN polimerasa, junto con muchos factores de transcripción, se une al promotor del gen. Esto forma un gran "complejo de pre-iniciación". Luego, el ADN se abre y la ARN polimerasa comienza a sintetizar el ARN. A veces, la polimerasa produce cadenas de ARN cortas que se liberan, hasta que finalmente logra hacer una cadena lo suficientemente larga para continuar.

Alargamiento de la Cadena en Eucariotas
Una vez que la ARN polimerasa ha iniciado la copia, se mueve a lo largo del ADN, desenrollándolo y añadiendo nucleótidos de ARN. A medida que el ARN se forma, las dos cadenas de ADN se vuelven a unir detrás de la polimerasa. Durante esta etapa, la ARN polimerasa puede hacer pausas en ciertos puntos, lo que a veces se usa para corregir errores o para regular la velocidad de la transcripción.
Final de la Copia en Eucariotas
La terminación de la transcripción es diferente para cada tipo de ARN polimerasa en eucariotas:
- Para Pol I: Necesita un factor especial para detenerse en sitios específicos del ADN.
- Para Pol II: Cuando la Pol II llega al final de un gen, reconoce una señal especial en el ARN que está copiando. Esto hace que el ARN se corte y se le añada una "cola" de muchas adeninas (llamada cola poli-A). Después de esto, la polimerasa se libera del ADN.
- Para Pol III: Se detiene de manera eficiente cuando encuentra una secuencia de timinas en el ADN.
Control de la Transcripción en Eucariotas
La regulación de la transcripción es muy importante en las células eucariotas. Permite que los genes se activen o desactiven según las necesidades de la célula. El ADN en eucariotas está muy organizado y enrollado alrededor de proteínas llamadas histonas, formando una estructura compacta llamada cromatina. Para que la transcripción ocurra, esta cromatina debe "abrirse" para que la ARN polimerasa pueda acceder al ADN.
Muchos factores, como los "potenciadores" y "silenciadores" (que son secuencias en el ADN), y proteínas activadoras o represoras, trabajan juntos para controlar cuándo y con qué intensidad se copia un gen. Este control asegura que cada célula produzca solo las proteínas que necesita en el momento adecuado.
Galería de imágenes
-
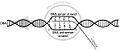
Esquema del proceso de transcripción de ARN. Se muestra la orientación antiparalela de las cadenas de un ADN y la producción de ARN por acción de la enzima ARN polimerasa.

