
Hipocampo (anatomía) para niños
El hipocampo es una parte muy importante de tu cerebro y el de otros mamíferos. Su nombre viene del griego y significa "caballo de mar", porque el anatomista Giulio Cesare Aranzio en el siglo XVI notó que se parecía a este animal.
El hipocampo se encuentra en la parte interna del lóbulo temporal de tu cerebro. Es una estructura con capas más sencillas de sustancia gris y forma parte de un grupo de estructuras llamado sistema límbico. Tenemos dos hipocampos, uno en cada lado del cerebro, como si fueran imágenes en un espejo. En los humanos y otros primates, tiene esa forma de caballito de mar, pero en otros animales puede ser diferente, como la de un plátano.
Aunque al principio se pensó que el hipocampo tenía que ver con el olfato, ahora sabemos que es clave para la memoria y para orientarnos en el espacio. Se ha estudiado mucho en animales como ratas y ratones, donde algunas de sus neuronas, llamadas "células de lugar", se activan cuando el animal está en un sitio específico. Estas células trabajan junto con otras que nos ayudan a saber hacia dónde vamos o a crear un "mapa" de nuestro entorno.
El hipocampo también es un lugar ideal para estudiar cómo las neuronas cambian y se fortalecen sus conexiones, un proceso llamado "potenciación a largo plazo" (LTP). Se cree que este proceso es fundamental para que el cerebro guarde los recuerdos.
Cuando el hipocampo se daña, por ejemplo, en enfermedades como el Alzheimer, o por falta de oxígeno o inflamación, las personas pueden tener problemas para formar nuevos recuerdos o para recordar cosas recientes.
Datos para niños Hipocampo |
||
|---|---|---|
 El hipocampo está localizado en la parte medial del lóbulo temporal del cerebro. En esta vista lateral del cerebro humano, el lóbulo frontal está a la izquierda, el lóbulo occipital a la derecha, y los lóbulos temporal y parietal han sido retirados en gran medida para revelar el hipocampo subyacente.
|
||
| Latín | [TA]: hippocampus | |
| TA | A14.1.09.321 | |
| Enlaces externos | ||
| NeuroLex ID | hippocampus | |
| NeuroNames | hippocampus | |
Contenido
¿Cómo se descubrió el hipocampo?
La primera vez que se describió el hipocampo fue por el anatomista Giulio Cesare Aranzi alrededor de 1587. Él lo comparó con un caballito de mar por su forma. Otros anatomistas de la época también lo dibujaron y le dieron nombres curiosos, como "gusano de seda" o "asta de carnero".
Más tarde, en 1742, se empezó a usar el nombre cornu Ammonis, que significa "cuerno de Amón", en honor al dios egipcio Amón. Este nombre todavía se usa para una parte del hipocampo.
Al principio, se pensaba que este órgano estaba relacionado con el olfato. Pero en los años 1900, el neurólogo ruso Vladímir Béjterev notó que el hipocampo tenía un papel importante en la memoria, al observar a un paciente con graves problemas para recordar. A pesar de esto, durante mucho tiempo se creyó que el hipocampo, como parte del sistema límbico, era responsable de las emociones.
¿Qué funciones tiene el hipocampo?
Con el tiempo, se han desarrollado tres ideas principales sobre lo que hace el hipocampo:
El hipocampo y la memoria
Los científicos están de acuerdo en que el hipocampo es crucial para formar nuevos recuerdos de lo que vivimos, como eventos o experiencias personales. También nos ayuda a detectar cosas nuevas, lugares o estímulos. Se considera parte de un sistema más grande en el lóbulo temporal que se encarga de la memoria declarativa, que es la que podemos expresar con palabras, como recordar hechos o eventos.
Si el hipocampo sufre daños graves, es muy difícil formar nuevos recuerdos (esto se llama amnesia anterógrada). A veces, también afecta a recuerdos formados antes del daño (amnesia retrógrada). Sin embargo, los recuerdos muy antiguos suelen conservarse, lo que sugiere que con el tiempo, los recuerdos se trasladan del hipocampo a otras partes del cerebro.
Es importante saber que el daño en el hipocampo no afecta a todos los tipos de memoria. Por ejemplo, no impide que aprendamos nuevas habilidades, como tocar un instrumento musical o resolver acertijos. Esto significa que estas habilidades dependen de otros tipos de memoria y de otras áreas del cerebro.
El hipocampo y la orientación espacial
Estudios en ratas y ratones han mostrado que las neuronas del hipocampo tienen "campos de lugar". Esto significa que se activan cuando el animal está en un lugar específico de su entorno. En humanos, también se han encontrado estas "células de lugar" en estudios con pacientes.
El descubrimiento de estas células llevó a la teoría de que el hipocampo funciona como un "mapa cognitivo", una especie de representación mental del espacio que nos rodea. Esta idea se apoya en que las personas con un hipocampo dañado a menudo se sienten perdidas y no recuerdan cómo llegar a un lugar.
Un estudio famoso con taxistas de Londres mostró que una parte de su hipocampo era más grande que en otras personas, y que los taxistas con más experiencia tenían un hipocampo más voluminoso. Esto sugiere que el uso constante de la memoria espacial puede hacer que esta parte del cerebro crezca.
¿Cómo se desarrolla el hipocampo?
Los hemisferios del cerebro empiezan a formarse en la quinta semana de desarrollo de un embrión. El hipocampo se forma a partir de una región llamada palio.
Antes se creía que el cerebro dejaba de producir nuevas neuronas después de la pubertad. Sin embargo, a finales del siglo XIX se descubrió que el hipocampo sigue generando nuevas neuronas durante toda la vida adulta. Esta producción de nuevas neuronas, llamada neurogénesis adulta, está muy relacionada con la capacidad de aprender y recordar.
¿Cómo funciona el hipocampo?
El hipocampo tiene dos "modos" principales de actividad, que se pueden ver en las ondas eléctricas del cerebro (EEG):
- Modo Theta: Aparece cuando estamos despiertos y activos (especialmente al movernos) o durante el sueño REM. En este modo, el EEG muestra ondas regulares. Las neuronas del hipocampo se activan de forma específica, y su actividad a menudo está relacionada con la ubicación del animal.
- Modo LIA (Actividad Irregular Lenta): Se observa durante el sueño profundo sin sueños y cuando estamos quietos, descansando o comiendo. En este modo, el EEG muestra ondas lentas y grandes, llamadas "ondas afiladas". Durante estas ondas, muchas neuronas se activan rápidamente.
Estas "ondas afiladas" parecen estar relacionadas con la memoria. Se ha visto que cuando las células de lugar del hipocampo se activan juntas durante una experiencia, tienden a activarse de nuevo de forma coordinada durante el sueño posterior. Esto se llama "reactivación" y se cree que ayuda a transferir los recuerdos del hipocampo a otras partes del cerebro para que se guarden a largo plazo.
Potenciación a largo plazo (LTP)
Desde hace mucho tiempo, los científicos han pensado que el cerebro guarda los recuerdos cambiando las conexiones entre las neuronas que se activan al mismo tiempo. En 1973, se descubrió en el hipocampo un fenómeno llamado "potenciación a largo plazo" (LTP). Es un cambio en la fuerza de las conexiones entre neuronas que dura mucho tiempo, incluso días o más, después de una activación fuerte y breve.
El hipocampo es un buen lugar para estudiar la LTP por sus neuronas bien organizadas. Se cree que la LTP es uno de los mecanismos principales por los que la memoria se almacena en el cerebro. Los medicamentos que interfieren con la LTP también afectan la memoria, especialmente la memoria espacial.
¿Qué le pasa al hipocampo con el tiempo o en ciertas condiciones?
El envejecimiento
Con la edad, incluso en un envejecimiento normal, algunas capacidades de la memoria pueden disminuir, como la memoria episódica (recuerdos de eventos) y la memoria de trabajo. Como el hipocampo es clave para la memoria, se investiga si estos cambios se deben a su deterioro. Algunos estudios han mostrado que el hipocampo puede reducir su tamaño en personas mayores, aunque no en todos. Se ha encontrado una relación entre el tamaño del hipocampo y el rendimiento de la memoria: las personas mayores con una memoria menos eficiente suelen tener un hipocampo más pequeño.
El estrés
El hipocampo es sensible al estrés prolongado. El estrés puede reducir la actividad de algunas neuronas, impedir la formación de nuevas neuronas y causar cambios en las conexiones de las neuronas. Se ha observado que personas que han vivido situaciones de estrés muy severo y prolongado pueden mostrar una reducción del tamaño del hipocampo. Estos efectos también se ven en el trastorno por estrés postraumático y pueden contribuir a cambios en el hipocampo en la esquizofrenia y la depresión. Algunos de estos efectos parecen ser reversibles si el estrés se detiene.
La epilepsia
El hipocampo es a menudo el origen de los ataques epilépticos. La esclerosis del hipocampo (un tipo de daño en el tejido) es común en la epilepsia del lóbulo temporal. No se sabe con certeza si la epilepsia es causada por problemas en el hipocampo o si el hipocampo se daña por los ataques repetitivos. El hipocampo es una de las partes del cerebro más excitables eléctricamente.
La esquizofrenia
En la esquizofrenia, se han encontrado varias anormalidades en la estructura del cerebro, incluyendo el hipocampo. Muchos estudios han mostrado que el hipocampo puede ser más pequeño en personas con esquizofrenia. Estos cambios probablemente se deben a alteraciones en el desarrollo, más que a un daño posterior. No está claro si estos cambios en el hipocampo causan los síntomas de la esquizofrenia, pero se cree que podrían afectar la forma en que el cerebro procesa la información.
¿Cómo ha evolucionado el hipocampo?
El hipocampo tiene un aspecto similar en todos los mamíferos, desde los monotremas hasta los humanos. Su tamaño en proporción al cuerpo aumenta en los primates, pero no tanto como otras partes del cerebro. Por eso, en roedores, el hipocampo ocupa una parte más grande de la corteza que en los primates.
Existe una relación general entre el tamaño del hipocampo y la memoria espacial. Las especies con mejor capacidad espacial suelen tener hipocampos más grandes. Esto también se ve en las diferencias entre machos y hembras en algunas especies: si hay grandes diferencias en la memoria espacial, también las hay en el volumen del hipocampo.
Aunque los no mamíferos no tienen un hipocampo idéntico, sí tienen una estructura cerebral similar, llamada palio medial, que se considera su equivalente. Se ha demostrado que estas estructuras similares al hipocampo también están involucradas en la orientación espacial en aves, reptiles y peces. Por ejemplo, las aves que esconden comida para el invierno tienen un hipocampo más grande y si se daña, tienen problemas para recordar dónde la guardaron.
Esto sugiere que el papel del hipocampo en la orientación espacial es muy antiguo en la evolución de los vertebrados.
Galería de imágenes
-

Hipocampo y fórnix comparados con un caballito de mar
-
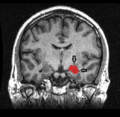
Hipocampo visto en una resonancia magnética
-

Vladímir Béjterev, neurólogo y psiquiatra soviético y probablemente el primero en asignar al hipocampo un papel en la formación de la memoria.
-
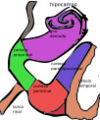
La formación hipocampal en relación con el resto de áreas corticales próximas del lóbulo temporal. El subículo está en el mismo color que el giro dentado y el hipocampo.
-

Patrones de disparo espacial de siete células de lugar registradas en la capa CA1 de rata. La rata recorre varios cientos de vueltas en el sentido de las agujas del reloj alrededor de una pista triangular, parando en el medio de cada uno de los lados para comer una pequeña porción de comida. Los puntos negros indican las posiciones de la cabeza de la rata. Los puntos coloreados indican los potenciales de acción, utilizando un color diferente para cada célula.
-
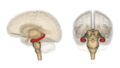
Hipocampo en rojo.
-
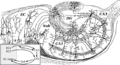
Circuito básico del hipocampo, según un dibujo de Santiago Ramón y Cajal. Siglas: DG: dentate gyrus (circunvolución dentada). Sub: subiculum. EC: corteza entorrinal.
-
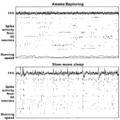
Ejemplos de electroencefalografía y actividad neural asociada a CA1 en hipocampo de rata en los modos theta (vigilia/conducta) y LIA (onda lenta, sueño). Cada punto muestra 20 segundos de datos, correspondiendo la gráfica superior al EEG de hipocampo, en el centro tramas de espigas de 40 células piramidales de la capa CA1 registradas simultáneamente (cada línea de la trama representa una célula diferente) y una gráfica de velocidad de carrera en el fondo. La gráfica superior presenta un período en el que la rata estaba buscando activamente piezas de comida distribuidas de forma dispersa. En la gráfica inferior, la rata estaba dormida.







.png)

Véase también
 En inglés: Hippocampus Facts for Kids
En inglés: Hippocampus Facts for Kids
- Subículo
- Indusium griseum
