
Intrón para niños
Un intrón es una parte del ADN que se encuentra dentro de un gen. Cuando el ADN se copia para formar una primera versión de ARN (llamada transcripción primaria), los intrones están presentes. Sin embargo, antes de que este ARN se use para crear proteínas, los intrones son eliminados. Las partes que quedan y que sí se usan para hacer proteínas se llaman exones.
Los intrones se encuentran en casi todos los seres vivos y también en los virus. La cantidad y el tamaño de los intrones pueden variar mucho entre diferentes especies. Por ejemplo, el pez globo tiene pocos intrones en su genoma, mientras que los mamíferos (como nosotros) y las plantas con flores suelen tener muchos.
La palabra intrón viene de "región intragénica", que significa una parte que está "dentro de un gen". A veces se les llama "secuencias que intervienen", pero este término puede referirse a otras partes del ácido nucleico que no terminan en el gen final.

Contenido
¿Quién descubrió los intrones?
Los intrones fueron descubiertos por los científicos Phillip Allen Sharp y Richard J. Roberts. Gracias a este importante hallazgo, recibieron el Premio Nobel de Fisiología o Medicina en 1993. El nombre "intrón" fue propuesto por el bioquímico estadounidense Walter Gilbert en 1978.
¿Para qué sirven los intrones?
Aunque por mucho tiempo se pensó que los intrones eran "ADN sin información", hoy sabemos que no es así. Los intrones son muy importantes para que los genes funcionen correctamente.
- Creación de diferentes proteínas: Los intrones pueden ayudar a que un solo gen produzca varios tipos de proteínas. Esto ocurre mediante un proceso llamado ayuste (o empalme) alternativo, donde se eligen diferentes combinaciones de exones para formar el ARN final.
- Información importante: Los intrones contienen pequeñas secuencias que son clave para que el proceso de ayuste sea eficiente y se realice de forma correcta.
- Ribozimas: Algunos intrones son especiales porque pueden "cortarse" a sí mismos del ARN sin ayuda de otras moléculas. A estas moléculas de ARN con capacidad de actuar como enzimas se les llama ribozimas. Este descubrimiento fue tan importante que Thomas R. Cech y Sidney Altman ganaron el Premio Nobel de Química en 1989 por ello.
Tipos de intrones
Existen cuatro tipos principales de intrones, que se clasifican según cómo son eliminados del ARN:
- Intrones del grupo I
- Intrones del grupo II
- Intrones del grupo III
- Intrones nucleares (también llamados espliceosomales o del grupo IV)
Los intrones de los grupos I, II y III se eliminan a sí mismos en un proceso llamado autoayuste. Son menos comunes que los intrones nucleares.
Los intrones del grupo II y III son muy parecidos en su estructura y cómo funcionan. A veces, los del grupo III se consideran parte del grupo II por su gran similitud.
- Intrones del grupo I: Se encuentran en algunos eucariotas (organismos con células complejas) y en los genes de las mitocondrias de los hongos. Para eliminarse, necesitan una molécula de guanosina.
- Intrones del grupo II y III: Se eliminan a sí mismos. Los del grupo II necesitan una adenina, y los del grupo III pueden necesitar un espliceosoma (una máquina molecular que corta y une el ARN). Durante su eliminación, forman una estructura especial en forma de lazo.
- Intrones del grupo IV: Estos se encuentran en los ARNt (ARN de transferencia) de los eucariotas. Son los únicos que se eliminan de una forma diferente: primero se cortan y luego se unen, en lugar de autoayustarse.
Funciones importantes de los intrones
Aunque los intrones no contienen las instrucciones para hacer proteínas, son fundamentales para controlar cómo se usan los genes.
- Regulación de genes: Algunos intrones, después de ser cortados, pueden convertirse en ARN no codificantes que tienen funciones importantes en la célula.
- Variedad de proteínas: El proceso de ayuste alternativo, donde se combinan diferentes exones, permite que un solo gen pueda producir muchas proteínas distintas. Esto es como tener un juego de piezas de construcción que se pueden armar de varias maneras para crear diferentes objetos.
- Protección del ADN: Las regiones de ADN que están activas a menudo forman estructuras que pueden ser dañadas. Los intrones ayudan a prevenir este daño, manteniendo el ADN más estable. Los estudios han mostrado que los genes con intrones tienen menos daño en el ADN que los genes sin intrones.
- Resistencia celular: La presencia de intrones también puede ayudar a las células a resistir la falta de nutrientes, regulando la producción de ciertas proteínas.
¿Cómo evolucionaron los intrones?
Existen dos ideas principales sobre cómo aparecieron y cambiaron los intrones a lo largo del tiempo:
- Modelo de intrones tempranos: Esta idea sugiere que los intrones eran muy comunes en los primeros seres vivos (ancestros de procariotas y eucariotas) y que se fueron perdiendo con la evolución. Se cree que los intrones ayudaron a combinar diferentes partes de los genes (exones), lo que facilitó la aparición de nuevos genes.
- Modelo de intrones tardíos: Este modelo propone que los intrones aparecieron después de que los procariotas y eucariotas se separaran. Se basa en que los intrones nucleares solo se han encontrado en eucariotas.
Los intrones pueden aparecer o desaparecer a lo largo de la evolución. Se han identificado miles de casos donde los intrones se han ganado o perdido en diferentes especies.
¿Cómo se ganan o pierden intrones?
Se conocen dos formas principales en que los intrones se pierden:
- Por un proceso que involucra una enzima llamada transcriptasa inversa.
- Por eliminaciones de partes del genoma.
Sin embargo, los mecanismos exactos de cómo se ganan nuevos intrones son más difíciles de entender y aún se investigan. Se han propuesto al menos siete formas en que un gen podría adquirir un nuevo intrón, como la transposición (cuando un intrón se mueve de un lugar a otro), la inserción de transposones (elementos genéticos móviles), o la "intronización" (cuando una parte del ADN que antes no era intrón se convierte en uno debido a cambios).
A pesar de estas ideas, los científicos siguen buscando pruebas directas de cómo surgen los nuevos intrones, ya que muchos de los intrones recién encontrados no parecen encajar perfectamente en los mecanismos propuestos. Esto sugiere que podría haber otros procesos aún por descubrir que generan intrones.
Galería de imágenes
-

Ilustración del proceso de ayuste desde pre-ARN a ARN.
-
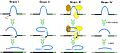
Ilustración que muestra la clasificación de los intrones según el método de ayuste.

Véase también
 En inglés: Intron Facts for Kids
En inglés: Intron Facts for Kids
