
Virus ADN para niños
Un virus ADN es un virus cuyo material genético principal es el ADN. Esto significa que, a diferencia de otros virus, no usa ARN como un paso intermedio para copiarse. Los virus que usan ARN, ya sea como su material genético o como un paso intermedio, se llaman virus ARN.
El ADN de un virus puede ser de una sola cadena (monocatenario) o de dos cadenas (bicatenario). Los virus con ADN de doble cadena son más comunes y variados. Para copiarse dentro de las células que infectan, estos virus necesitan una enzima especial llamada ADN polimerasa. A menudo, los virus con ADN de una sola cadena se convierten en ADN de doble cadena una vez que están dentro de la célula infectada.
¿Qué es un Virus ADN?
Un virus ADN es un tipo de virus que guarda su información genética en una molécula llamada ADN. Esta molécula es como un manual de instrucciones que el virus usa para hacer más copias de sí mismo. A diferencia de los virus ARN, los virus ADN no necesitan usar ARN como un paso intermedio para replicarse.
¿Cómo se clasifican los Virus ADN?
Los científicos usan un sistema llamado clasificación de Baltimore para organizar los virus. Este sistema los agrupa según cómo producen su ARN mensajero (ARNm), que es una molécula que lleva las instrucciones del ADN para fabricar proteínas. Esta clasificación se usa junto con la forma tradicional de clasificar virus, que se basa en su historia evolutiva.
Los virus ADN se dividen en tres grupos principales dentro de la clasificación de Baltimore:
- Grupo I: Virus ADN bicatenario (con ADN de doble cadena).
- Grupo II: Virus ADN monocatenario (con ADN de una sola cadena).
- Grupo VII: Virus ADN bicatenario retrotranscrito (con ADN de doble cadena que usa un paso de ARN para copiarse).
Aunque la clasificación de Baltimore se enfoca en cómo se produce el ARNm, los virus de cada grupo también suelen compartir la forma en que se copian. Es importante saber que los virus en un mismo grupo de Baltimore no siempre están relacionados genéticamente o tienen la misma forma.
¿Cómo se copian los Virus ADN bicatenarios (Grupo I)?
Los virus del Grupo I tienen un genoma de ADN de doble cadena. Para producir su ARNm, siguen un proceso de tres pasos:
- Primero, un grupo de proteínas se une al ADN del virus, lo que permite que una enzima de la célula huésped, llamada ARN polimerasa, se una también.
- Segundo, la ARN polimerasa usa una de las cadenas del ADN como molde para crear el ARNm.
- Tercero, la ARN polimerasa deja de producir ARNm cuando llega a una señal específica en el ADN.
Estos virus usan diferentes maneras para copiar su genoma:
- Replicación bidireccional: Es una forma común de copiar ADN. El ADN circular se abre, creando dos puntos donde la copia comienza y avanza en direcciones opuestas hasta que todo el ADN se ha copiado.
- Círculo rodante: En este método, el ADN circular se copia continuamente, produciendo largas cadenas lineales que luego se cortan en genomas individuales.
- Desplazamiento de cadena: Una cadena de ADN se copia primero, y luego la otra cadena se copia usando la primera como molde.
- Transposición replicativa: El genoma viral se copia y se inserta en otra parte del ADN de la célula huésped.
Algunos virus ADN bicatenarios se copian en el núcleo de la célula y dependen mucho de la maquinaria de la célula. Otros se copian en el citoplasma (la parte líquida de la célula) y han desarrollado sus propias herramientas para hacerlo.
¿Cómo se copian los Virus ADN monocatenarios (Grupo II)?
Los virus del Grupo II tienen un genoma de ADN de una sola cadena. Cuando entran en una célula, su ADN de una sola cadena se convierte primero en una forma de doble cadena gracias a una ADN polimerasa. Luego, el ARNm se produce a partir de esta forma de doble cadena.
La mayoría de estos virus tienen genomas circulares que se copian mediante el método de círculo rodante. Una enzima especial corta una de las cadenas, permitiendo que la ADN polimerasa use la otra cadena como molde para copiar el ADN. El nuevo ADN de una sola cadena puede ser empaquetado en nuevas partículas virales o convertirse de nuevo en doble cadena para seguir copiándose.
Los parvovirus, que son un tipo de virus ADN monocatenario, tienen genomas lineales y usan un método llamado horquilla rodante. Sus genomas tienen estructuras en forma de horquilla en los extremos que se abren y cierran, permitiendo que el ADN se copie de forma continua.
Casi todos los virus ADN monocatenarios tienen genomas de "sentido positivo", pero hay algunas excepciones.
¿Cómo se copian los Virus ADN bicatenarios retrotranscritos (Grupo VII)?
Los virus del Grupo VII tienen un genoma de ADN de doble cadena que se copia usando un paso intermedio de ARN. Antes de producir ARNm, cualquier espacio en su ADN se repara para formar un genoma completo.
Estos virus producen ARNm de manera similar a los virus ADN bicatenarios. Sin embargo, para copiar su genoma, usan una enzima llamada transcriptasa inversa que convierte el ARN en ADN. La ARN polimerasa II de la célula huésped crea cadenas de ARN a partir del genoma viral en el citoplasma, y luego el genoma de ADN se copia a partir de estas cadenas de ARN. Después de copiarse, el nuevo genoma de ADN puede ser empaquetado en nuevas partículas virales o enviado al núcleo para seguir produciendo ARNm.
¿De dónde vienen los Virus ADN?
Los virus ADN son muy antiguos y se encuentran en muchos tipos de seres vivos. Los científicos creen que tienen orígenes muy diversos y que han evolucionado de diferentes maneras a lo largo del tiempo.
El Comité Internacional de Taxonomía de Virus (ICTV) ha identificado varios grupos grandes de virus ADN, llamados dominios, que comparten características especiales:
- Duplodnaviria: Incluye virus con ADN de doble cadena y una proteína específica en su cubierta. Se cree que surgieron hace mucho tiempo, incluso antes del último ancestro común de todos los seres vivos.
- Adnaviria: Son virus de arqueas (un tipo de microorganismo) con ADN de doble cadena y forma filamentosa. También se piensa que son muy antiguos.
- Varidnaviria: Contiene virus con ADN de doble cadena y una proteína especial en su cubierta. Incluye virus como los adenovirus y los virus gigantes.
- Monodnaviria: Agrupa a casi todos los virus con ADN de una sola cadena, además de algunos con ADN de doble cadena que evolucionaron de ellos. Se cree que surgieron cuando pequeños fragmentos de ADN se unieron a virus de ARN.
- Singelaviria: Son virus con ADN de doble cadena que solo infectan procariotas (organismos sin núcleo definido).
- Telodnaviria: Incluye virus con ADN de doble cadena de forma filamentosa u ovoide.
- Riboviria: Este dominio es especial porque incluye a todos los virus de ARN y a los virus que usan la transcriptasa inversa (como los virus ADN bicatenarios retrotranscritos), aunque tengan ADN. Se caracterizan por tener enzimas muy antiguas que copian ácidos nucleicos.
Además de estos dominios, existen algunas familias de virus ADN que aún no han sido clasificadas en ninguno de estos grupos grandes. Se cree que algunos de estos virus inusuales de arqueas podrían estar relacionados con elementos genéticos móviles (pequeños fragmentos de ADN que pueden moverse dentro de un genoma) en lugar de con otros virus.

Galería de imágenes
-
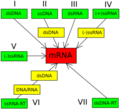
Clasificación de Baltimore de los virus, basada en la obtención del ARNm a partir del genoma del virus. Los Grupos I, II y VII son virus ADN.
Véase también
 En inglés: DNA virus Facts for Kids
En inglés: DNA virus Facts for Kids
