
Fragmentación de hábitat para niños
La fragmentación de hábitat es un proceso importante que cambia el ambiente y afecta la evolución y la biología de la conservación. Imagina un gran espacio natural, como un bosque o una pradera. La fragmentación ocurre cuando ese espacio se divide en pedazos más pequeños. Estos pedazos quedan separados por áreas que ya no son naturales, como ciudades, carreteras o campos de cultivo.
A medida que el paisaje se fragmenta, los pedazos de hábitat se acercan a estas nuevas áreas. Los límites entre el hábitat natural y lo que lo rodea se llaman bordes. La extensión de estos bordes depende de cómo sea el área que rodea el hábitat. La fragmentación es un proceso que va más allá de la interacción entre los pedazos de hábitat y las áreas que los separan. Para que realmente haya fragmentación, el paisaje debe perder su continuidad. Esto significa que debemos estudiar este fenómeno a una escala más grande, regional. La conexión de un hábitat depende de dónde estén los fragmentos y de las características del ambiente que los rodea.
Aunque la fragmentación puede ocurrir por causas naturales, como incendios o cambios geológicos, la razón más común y extendida es la actividad humana. Los seres humanos usamos cada vez más la tierra, construyendo y cultivando. Esto, junto con las graves amenazas a la biodiversidad (la variedad de vida en la Tierra), ha convertido la modificación y fragmentación del paisaje en uno de los problemas más urgentes en la ecología de la conservación.
El estudio de la fragmentación comenzó con la teoría de la biogeografía de islas. Esta teoría se enfoca en cómo el tamaño y el aislamiento de un lugar afectan la cantidad de especies que viven allí. Se decía que el número de especies en una isla (o un pedazo de hábitat) dependía del equilibrio entre la llegada de nuevas especies (colonización) y la desaparición de otras (extinción). Las islas más pequeñas y aisladas tendrían menos especies. Sin embargo, esta teoría solo considera algunos aspectos, lo que llevó a buscar otros enfoques.
Bascompte y Solé sugieren que la mejor manera de estudiar la fragmentación es a través de modelos de metapoblaciones. Estos modelos describen cómo se comportan grupos de poblaciones pequeñas que viven en áreas separadas. Este enfoque es más dinámico y diferente de la teoría de islas. Considera el paisaje como una red de pequeños pedazos de hábitat que cambian de tamaño con el tiempo. Destaca que el movimiento de especies entre estos pedazos es clave para su supervivencia. Si no hay un área fija, una dispersión inadecuada podría llevar a la extinción de especies, tanto a nivel local como regional.
Contenido
- ¿Qué es la pérdida de hábitat y el umbral de extinción?
- ¿Cómo afecta la fragmentación a la genética de las especies?
- ¿Cuáles son los efectos de la fragmentación en la biodiversidad?
- ¿Cómo se detecta el impacto de la fragmentación?
- Fragmentación de hábitat y cambio climático
- ¿Qué podemos hacer para conservar los hábitats?
- Galería de imágenes
- Véase también
¿Qué es la pérdida de hábitat y el umbral de extinción?

El efecto más claro de la fragmentación es la pérdida de hábitat natural. Por eso, muchos investigadores miden el impacto simplemente por la cantidad de hábitat que queda. Sin embargo, la fragmentación puede ocurrir de muchas maneras, creando patrones diferentes. La pérdida de hábitat siempre va de la mano con la fragmentación, pero son fenómenos distintos. La fragmentación por sí misma tiene impactos importantes en la biodiversidad, incluso sin considerar solo la pérdida de hábitat.
Para entender la fragmentación y sus consecuencias, se han creado mapas con modelos aleatorios. Estos muestran que el tamaño y el aislamiento de los pedazos de hábitat no cambian de forma lineal con la cantidad de hábitat perdido. Esto significa que hay ciertos puntos, llamados umbrales, donde pequeñas pérdidas de hábitat pueden acelerar drásticamente la disminución del tamaño de los pedazos y aumentar su aislamiento. Esto hace que las poblaciones que quedan sean inviables. Pequeñas pérdidas de hábitat cerca de estos umbrales pueden reducir mucho la probabilidad de supervivencia de las metapoblaciones.
Estos umbrales en el proceso físico de fragmentación son muy importantes para la conservación de la biodiversidad. Diferentes especies desaparecen en distintos puntos a lo largo de la destrucción del hábitat. El umbral de extinción es la cantidad mínima de hábitat que una población necesita para sobrevivir. Esto sugiere que los umbrales de extinción dependen de cómo varía la fragmentación, pero también de las características de cada especie y del ambiente en el que viven. Por ejemplo, With y King descubrieron que el umbral de extinción puede variar desde un 5% hasta un 90% de hábitat destruido, según la especie y cómo estén distribuidos los fragmentos.
La ciencia ambiental ha establecido medidas para la destrucción de la biodiversidad causada por problemas como la deforestación, el cambio climático, la introducción de especies invasoras o el uso excesivo de los recursos naturales. Esto se conoce como la Deuda de extinción.
¿Cómo afecta la fragmentación a la genética de las especies?
La fragmentación crea poblaciones pequeñas, separadas y aisladas. Esto puede afectar la genética de las especies. Se espera que la fragmentación reduzca la variación genética y aumente las diferencias entre poblaciones. Esto se debe a que hay más "deriva genética" (cambios aleatorios en los genes) y "endogamia" (reproducción entre individuos emparentados), y menos "flujo genético" (intercambio de genes) entre las poblaciones que quedan.
La fragmentación puede disminuir la variedad genética de una población de dos maneras. Primero, cuando una población grande se fragmenta, solo una pequeña parte de sus genes originales se mantiene. Esto se llama "cuello de botella genético". Los efectos inmediatos dependen del tamaño de las poblaciones que quedan y de la variedad genética que tenían al principio.
Después del cuello de botella, si las poblaciones siguen siendo pequeñas y aisladas por varias generaciones, la variedad genética disminuirá. Esto puede llevar a la acumulación de genes dañinos y a una menor capacidad de reproducción, lo que podría llevar a la extinción de la población. La pérdida de variedad genética puede reducir la capacidad de las poblaciones para adaptarse a futuros cambios ambientales, aumentando su riesgo de extinción.
Sin embargo, los estudios no siempre muestran los mismos resultados. No todos los casos de fragmentación en plantas han llevado a una pérdida genética. Esto puede explicarse por las características de cada planta (como su tiempo de vida o cómo se reproducen), que pueden hacerlas más o menos vulnerables. Además, el tiempo que ha pasado desde la fragmentación y otros procesos ecológicos pueden influir en la conexión genética y demográfica.
¿Cuáles son los efectos de la fragmentación en la biodiversidad?
La fragmentación crea paisajes y pedazos de hábitat con características específicas. Estas características se pueden agrupar en cinco categorías:
- (1) el tamaño del fragmento
- (2) el efecto de borde
- (3) la forma del fragmento
- (4) el aislamiento del fragmento
- (5) la estructura del área que rodea el fragmento (la matriz)
Tamaño del fragmento
Algunos pedazos de hábitat pueden ser demasiado pequeños para que ciertas poblaciones sobrevivan, lo que reduce mucho su tamaño. Esto puede causar extinciones locales, y por eso, los pedazos pequeños suelen tener menos especies. Esto es común en animales grandes. Sin embargo, algunos estudios han encontrado relaciones diferentes entre el tamaño del animal y la probabilidad de extinción.
El hecho de que el tamaño del fragmento limite el tamaño de la población hace que las especies sean más vulnerables a las extinciones locales. Esto las expone a la pérdida genética y a problemas por endogamia. Los motivos por los que el tamaño del área afecta la vulnerabilidad incluyen: cambios ambientales aleatorios, cambios demográficos aleatorios, desastres naturales y reducción de la variedad genética. Aunque es útil distinguirlos, rara vez actúan de forma independiente.
Además, el hábitat no es uniforme. Los fragmentos individuales pueden no contener todos los tipos de microhábitats que había en el hábitat original. Las especies que necesitan varios microhábitats son especialmente vulnerables, ya que deben moverse entre fragmentos para satisfacer sus necesidades, lo cual a veces es imposible.
También se ha demostrado que los fragmentos pequeños afectan varios aspectos de las poblaciones y comunidades. Por ejemplo, reducen la longitud de las cadenas alimentarias, el número de especies especialistas y de gran tamaño, y alteran las interacciones entre especies. La fragmentación también afecta negativamente el éxito reproductivo, la dispersión, la tasa de depredación y el comportamiento de los animales al buscar alimento.
Efectos de borde

Los efectos de borde son cambios en las condiciones físicas y biológicas que ocurren en los límites artificiales y abruptos de los fragmentos de hábitat. Los efectos no vivos (abióticos) se refieren a cambios en la luz solar, humedad, temperatura, velocidad del viento y nutrientes del suelo. Los efectos vivos (bióticos) son cambios en la composición de especies o en los patrones de competencia, depredación y parasitismo. La profundidad de estos efectos en el fragmento puede variar mucho, desde decenas de metros hasta varios kilómetros, debido a factores como el contraste con el área circundante, la velocidad del viento, la temperatura y la presencia de especies invasoras. Además, los bordes cercanos a áreas urbanas sufren modificaciones por la actividad humana.
El hábitat de los bordes puede cambiar la naturaleza de las interacciones y, por lo tanto, modificar procesos ecológicos. La estructura, composición y diversidad de las comunidades se ven afectadas de diversas maneras en las zonas de borde. Por ejemplo, en las comunidades de invertebrados, la riqueza de especies (cantidad de especies) suele disminuir a medida que se avanza hacia el centro del fragmento. Una explicación común es que la mezcla de especies de ambos lados supera en número a las especies perdidas. Sin embargo, esto no siempre es así; algunos estudios han encontrado que muchas especies evitan los bordes, y que el área circundante no siempre tiene suficientes especies para compensar las pérdidas. Curiosamente, algunos animales parecen preferir los bordes para reproducirse, aunque las tasas de mortalidad allí sean mucho más altas. En esos casos, se les llama trampas ecológicas.
Forma del fragmento

La forma de un fragmento se define por la relación entre su área y su perímetro. Esto, a su vez, determina la cantidad de hábitat que permanece intacto en el centro (el núcleo). Por eso, los fragmentos con formas complejas tienen una mayor proporción de su área como borde. Además, las formas muy irregulares pueden crear divisiones adicionales en el núcleo del fragmento, generando varios núcleos separados por hábitat de borde. Sin embargo, el efecto de la forma no es independiente del tamaño. El impacto de los efectos de borde aumenta a medida que el fragmento se hace más pequeño, ya que cualquier desviación de una forma circular reduce significativamente la cantidad de hábitat interior.
El patrón más consistente relacionado con la forma de los fragmentos es que aquellos con formas complejas son colonizados con más frecuencia. Esto se debe a que tienen una mayor proporción de borde, lo que aumenta la probabilidad de que un animal en movimiento encuentre el fragmento. Esta probabilidad también puede depender de la orientación del fragmento. Si las especies se mueven en una dirección predecible (como en migraciones), los fragmentos largos y estrechos son más propensos a ser colonizados, pero solo si están orientados perpendicularmente a la dirección del movimiento. Sin embargo, así como aumenta la colonización, también lo hace la salida de individuos. El resultado es una menor probabilidad de que las poblaciones permanezcan en estos fragmentos.
Aislamiento del fragmento

Una de las consecuencias más claras de la fragmentación es el aislamiento de los pedazos de hábitat en el tiempo y el espacio. Este aislamiento se entiende como una medida de cómo está configurado el hábitat. Sin embargo, es más preciso interpretarlo como una medida de la cantidad de hábitat que se ha eliminado.
El aislamiento interrumpe los patrones de distribución de las especies y obliga a los individuos a moverse a través del área que rodea el hábitat (la matriz). Dado que la matriz puede dificultar la dispersión de algunas especies más que de otras, y que las especies difieren en su disposición a cruzarla, se han encontrado resultados contradictorios. En insectos, por ejemplo, la diferencia genética entre poblaciones se ha relacionado claramente con el aislamiento en algunos estudios, pero no en otros.
En resumen, las interrupciones pueden dividir las metapoblaciones, disminuyendo el tamaño total de la población. Esto aumenta la probabilidad de extinciones locales y evita que nuevas colonizaciones restablezcan poblaciones que ya se extinguieron. Por eso, una de las medidas de conservación más intuitivas es la creación de corredores biológicos.
Estructura de la matriz
La idea de que los paisajes modificados son como "islas" ha influido mucho en la literatura sobre fragmentación. Sin embargo, la teoría tradicional de Wilson y MacArthur no reconoce que los efectos de borde pueden alterar las características del fragmento, y que la matriz (el área que rodea los fragmentos) puede no ser completamente inhóspita para la fauna nativa. De hecho, cada vez hay más pruebas que sugieren que las características de la matriz son cruciales para determinar la abundancia y composición de especies dentro de los fragmentos. Afecta principalmente las características del borde y la dispersión y conexión entre los pedazos de hábitat.
La dispersión entre fragmentos es esencial para la supervivencia a largo plazo de las metapoblaciones, y depende en gran parte de las características de la matriz. Esta actúa como un filtro selectivo para el movimiento de propágulos (semillas, esporas, etc.) y animales, afectando la composición de especies en los fragmentos. Su estructura, por ejemplo, puede alterar la dinámica de colonización-extinción y provocar cambios en la densidad poblacional, la estructura de los fragmentos y las interacciones entre especies. Además, la matriz puede influir en la naturaleza y magnitud de los efectos de borde. Los fragmentos y las matrices muy diferentes entre sí generan efectos de borde más severos y son más difíciles de cruzar para los animales.
Por otro lado, los restos de hábitat nativo no son necesariamente las únicas partes del paisaje que proporcionan recursos importantes para la supervivencia de las especies. Algunas especies pueden compensar la pérdida de hábitat utilizando los recursos presentes en la matriz. Mediante modelos de simulación, se ha demostrado que las especies generalistas (es decir, las que pueden usar recursos de la matriz) alcanzan tamaños de población mayores que las especies especialistas. Asimismo, modelos similares muestran que al mejorar la calidad de la matriz (es decir, aumentando los recursos que aporta al sistema) se incrementa la densidad de población dentro de los fragmentos. De este modo, la capacidad de las especies para utilizar recursos de la matriz puede alterar la intensidad de los efectos de la fragmentación del hábitat.
¿Cómo se detecta el impacto de la fragmentación?
Los estudios sobre el impacto de la fragmentación pueden variar mucho en sus resultados, dependiendo de lo que se incluya en la investigación. Esto explica por qué hay tantos ejemplos aparentemente contradictorios en la literatura sobre fragmentación de hábitat, que no pueden explicarse solo por diferencias en los ambientes o métodos. Estas diferencias resaltan la gran cantidad de factores que pueden ocultar o realzar los efectos de la fragmentación.
Estos factores pueden ser los mecanismos que explican los patrones observados en las especies, o las características de las especies que las hacen más o menos vulnerables a la fragmentación. En general, se cree que ciertos atributos aumentan la vulnerabilidad de las especies a la fragmentación, como tener cuerpos grandes, poca movilidad o dispersión, poca tolerancia a la matriz, dependencia de interacciones de ayuda mutua y niveles tróficos altos (estar en la cima de la cadena alimentaria). De hecho, se ha demostrado que la extinción de especies ocurre de forma consistente y secuencial a medida que el área del fragmento disminuye, lo que indica una vulnerabilidad diferente entre las especies. Así, estudiar los impactos de la fragmentación en especies con umbrales de extinción diferentes puede generar resultados que parecen contradictorios.
Además, las especies tienen un conjunto de características que están muy relacionadas y que pueden interactuar entre sí, y con las de otras especies, modificando o aumentando los impactos de la fragmentación. Un ejemplo es la interacción planta-polinizador, ya que se ha documentado que la pérdida y fragmentación del hábitat pueden alterar significativamente las comunidades de polinizadores e interrumpir esta interacción.
Por otro lado, hay un gran componente temporal en cómo las especies responden a la fragmentación. La mayoría de los estudios sobre paisajes fragmentados por la acción humana se han realizado en menos de cien años después de la fragmentación, por lo que los efectos a largo plazo son relativamente desconocidos. A corto plazo, se sabe que los organismos que sobreviven a la pérdida inmediata de hábitat se concentran en superficies mucho más pequeñas, lo que genera efectos de hacinamiento y aumenta la densidad de población y de especies. Sin embargo, los fragmentos no pueden soportar a todos los individuos y especies a largo plazo, lo que provoca una reducción progresiva en la abundancia de especies con el tiempo. Estas reducciones se llaman comúnmente deudas de extinción, y ocurren a medio y largo plazo, describiendo un retraso temporal entre la pérdida del hábitat y el eventual colapso de las poblaciones. La deuda de extinción se ilustra con correlaciones negativas entre la riqueza de especies y la edad del fragmento. Sin embargo, varios estudios han encontrado lo contrario en fragmentos de edad intermedia y prolongada, lo que sugiere que después del impacto, las comunidades podrían alcanzar nuevos "equilibrios".
Otro factor que puede influir en los impactos de la fragmentación es la biogeografía y la historia de los ambientes. La sensibilidad de las especies es diferente según los distintos biomas, siendo particularmente baja en las zonas templadas del hemisferio norte. Una posible explicación es la del filtro de extinción, que sugiere que la fragmentación en esos biomas es lo suficientemente antigua como para que las especies más vulnerables ya se hayan extinguido. También se cree que las comunidades de regiones templadas son más resistentes a los efectos de la fragmentación del hábitat que las comunidades de regiones tropicales, porque las primeras están más ampliamente distribuidas, ocurren en mayores densidades y tienen mayor capacidad de dispersión que sus contrapartes tropicales.
Finalmente, hay que tener en cuenta que la fragmentación puede tener efectos positivos para la biodiversidad, aunque estos pueden quedar ocultos por los efectos negativos de la pérdida de hábitat. Por ejemplo, Huffaker realizó experimentos que sugieren que dividir la misma cantidad de hábitat en muchos pedazos pequeños puede favorecer la supervivencia de sistemas depredador-presa. Él planteó la hipótesis de que la subdivisión proporciona refugios temporales para las presas, donde estas pueden aumentar en número y dispersarse a otros sitios antes de que el depredador o parásito las encuentre; una idea que puede aplicarse al proceso de fragmentación.
Fragmentación de hábitat y cambio climático
Modelos recientes han indicado que la pérdida y fragmentación del hábitat pueden aumentar la vulnerabilidad de las especies al aumento de la temperatura y a la frecuencia de eventos climáticos extremos que provienen del cambio climático global. Esto se debe a que cuando las poblaciones están aisladas, su área de expansión está limitada. Esta limitación no permite que las especies escapen de las condiciones desfavorables, una estrategia de supervivencia típica de ciertos insectos. Como consecuencia de esta estrategia, recientemente en Europa se ha extendido la distribución en el límite norte de varias especies de mariposas debido al calentamiento global; e incluso se espera una mayor expansión en el futuro. Este cambio en el rango de distribución de las especies es, en realidad, el resultado de la extinción de poblaciones en los límites cálidos de su área de distribución, y la colonización y crecimiento de las poblaciones en los límites más frescos. Se cree que esta dinámica es más rápida en regiones donde la estructura del paisaje permite la dispersión, en comparación con regiones donde el paisaje está fragmentado. Es decir, que a partir de la interacción asumida entre el cambio climático y la fragmentación, se generarían grandes tasas de expansión en paisajes que tengan alta cohesión espacial; aunque no existen estudios que apoyen esta predicción.
Un factor importante a considerar es que el cambio climático debe ocurrir a una velocidad que permita la expansión y colonización de nuevos ambientes antes de que la población que proporciona los propágulos se extinga. Esta coordinación depende a su vez de las características de las especies, como puede ser el rango de dispersión de propágulos. Sin embargo, si las especies necesitan largos períodos de tiempo o numerosos eventos de colonización para establecer nuevos asentamientos, la tasa de expansión será mucho menor de lo esperado según la capacidad de dispersión de la especie. Asimismo, el proceso de extinción-recolonización causa una distribución menos efectiva a través del hábitat, y la mortalidad de los propágulos es alta, lo que extiende el tiempo de recuperación de las metapoblaciones. En conclusión, se puede decir que la fragmentación del hábitat multiplica el impacto del cambio climático.
¿Qué podemos hacer para conservar los hábitats?
Como se mencionó antes, la modificación y fragmentación del paisaje son temas muy serios y urgentes en la ecología de la conservación. Por eso, hay muchos estudios teóricos y observacionales al respecto. A partir de ellos, se han propuesto diferentes medidas, muchas de ellas complementarias, para reducir o evitar las consecuencias negativas de la fragmentación.
Una de las medidas más conocidas es la creación de reservas naturales. Idealmente, el propósito de estas reservas es mantener un conjunto complejo de procesos físicos, ecológicos, genéticos, de comportamiento y evolutivos, y las poblaciones compatibles que participan en ellos. Ante tanta complejidad, las dificultades básicas en la planificación son: la cantidad, forma y cercanía de las reservas. La solución varía según la escala del esfuerzo de conservación. Al intentar resolver estas dificultades, la teoría de la fragmentación sentó las bases del acrónimo SLOSS (una grande o varias pequeñas), generando grandes debates. A pesar del intenso debate, se considera que las reservas de gran tamaño son mejores para la supervivencia a largo plazo de las especies, ya que mantienen poblaciones más grandes y el hábitat está menos expuesto a la degradación con el tiempo.
Por otro lado, uno de los métodos claros para frenar y compensar el avance de las extinciones locales es la recolonización artificial. Para ello, los recursos genéticos son una herramienta muy útil. Estas intervenciones no solo son necesarias para evitar extinciones, sino que también permiten asegurar la permanencia de un número suficiente de poblaciones locales y preservar la variedad genética global. Existen dos estrategias básicas: recolonizar con animales de una única población grande, o con una mezcla de animales de varias poblaciones. Esta última puede restablecer la variedad a nivel local, pero como las poblaciones introducidas suelen ser pequeñas, esta variación puede perderse rápidamente por deriva genética. Sin embargo, incluso una infusión temporal de variedad genética puede ser beneficiosa. Otros factores a considerar son la depresión por "exogamia" (la ruptura de genotipos adaptados por recombinación) y la depresión por "endogamia" (debido al pequeño tamaño de la población). Estos podrían poner en peligro la población introducida en las primeras generaciones hasta que se seleccionen y establezcan genotipos adaptados nuevamente.
A pesar de los riesgos en las primeras generaciones, la introducción de poblaciones mezcladas permite que los alelos dañinos se eliminen por selección natural. Por otro lado, la introducción de individuos de una única población no lo permite, por lo tanto, cualquier característica puede fijarse por endogamia, poniendo en riesgo la población. Esto sugiere que es preferible liberar una mezcla de individuos, idealmente cerca de la capacidad de carga del sistema. Pero el proceso de recolonización continúa, ya que la población liberada debe ser monitoreada genética y demográficamente en las primeras generaciones para detectar posibles problemas por exogamia o endogamia.
Desde otra perspectiva, se ha propuesto reducir los efectos negativos del aislamiento de los fragmentos manteniendo la conectividad del paisaje mediante matrices suaves, puntos clave, y principalmente corredores biológicos. En el contexto ecológico de la fragmentación, el término corredor se refiere a un elemento lineal del paisaje compuesto por vegetación nativa que une pedazos de vegetación nativa similar. Estas conexiones facilitan el movimiento de plantas y animales entre los diferentes pedazos, permitiendo que existan más especies y/o permanezcan por más tiempo de lo esperado según el tamaño de los mismos; es decir, que se incrementa el tamaño efectivo de los fragmentos.
La utilidad de los corredores como hábitat o posibles rutas de dispersión varía según las especies. En el caso de los animales, depende de sus patrones de alimentación, tamaño corporal, tamaño del territorio, grado de especialización en la dieta, movilidad y comportamiento social. Además, la percepción y el uso de los corredores por los animales difieren según sus dimensiones físicas y el contexto del paisaje.
Sin embargo, no todas las consecuencias de los corredores son positivas. Por ejemplo, pueden facilitar la expansión de la distribución de especies exóticas invasoras. Asimismo, mediante simulaciones se ha demostrado que cuando la interacción huésped-parásito es importante en la dinámica del huésped, el aumento de la conectividad del paisaje promueve la transmisión del parásito, aumentando la probabilidad de extinción de la metapoblación. Cabe aclarar que modelos posteriores han indicado que los beneficios de los corredores suelen compensar o superar los riesgos de la transmisión de parásitos. Por lo tanto, a pesar de algunos riesgos ecológicos, al aumentar la conectividad del paisaje es más probable encontrar efectos positivos que negativos en las especies nativas y los procesos ecológicos.
Por último, cabe mencionar que se ha recomendado dejar de lado el enfoque de conservación a nivel de especies, para centrarse en las condiciones en las que se desarrollan las comunidades. Estas condiciones deben permitir que las poblaciones respondan a grandes cambios y se adapten genéticamente a ellos. Para ello, hay que tener en cuenta que los factores que influyen en las poblaciones son numerosos, y por lo tanto, los planes de conservación que solo se basen en el área a preservar probablemente tendrán un éxito limitado.
Galería de imágenes
-
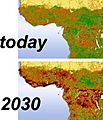
Fragmentación y destrucción del hábitat de los grandes simios en África Central entre 2002 y 2030. Información de GLOBIO y el proyecto GRASP.
-

Fragmento de bosque aislado experimentalmente en el Amazonas central (fotografiado por R.O. Bierregaard).


Véase también
 En inglés: Habitat fragmentation Facts for Kids
En inglés: Habitat fragmentation Facts for Kids
- Perturbación (ecología)
- Biodiversidad
- Destrucción de hábitat
- Ecosistema
- Hábitat
- Deuda de extinción
