
Evolución del ojo para niños
La capacidad de ver es increíble, ¿verdad? Nuestros ojos nos permiten explorar el mundo, ver colores y formas. Pero, ¿alguna vez te has preguntado cómo evolucionaron los ojos? Es una historia fascinante de cómo los seres vivos se adaptaron a la luz que los rodea.
La luz, en sus diferentes cantidades y tipos, ha sido una de las fuerzas más importantes que impulsaron la evolución del ojo. Esto se debe a que los distintos tipos de ojos son, en realidad, formas en que los organismos se adaptaron a su entorno.
Hay una gran variedad de formas y partes (como lentes y células que detectan la luz) en los ojos de los animales. Esto podría hacernos pensar que el sistema visual apareció muchas veces de forma independiente en diferentes grupos. Sin embargo, hay un gen llamado Pax6 que es muy similar en animales muy distintos y que es clave en la formación del ojo. Esto nos hace pensar que, quizás, todos los ojos tienen un origen común.
Contenido
Tipos de Ojos: ¿Cómo Ven los Animales?

Existen dos grandes tipos de ojos: los ojos compuestos y los ojos simples. Si observamos cómo se distribuyen estos tipos de ojos en el árbol de la vida, vemos que no hay un patrón único. Muchos grupos de animales tienen más de un tipo de ojo, y a veces, un tipo de ojo se repite en grupos muy diferentes. Incluso, algunos animales han perdido la vista por completo porque viven en la oscuridad o son parásitos.
Ojos Sencillos: Detectando Luz y Oscuridad
Los ojos más simples son solo un grupo de células que detectan la luz. Estos ojos permiten al animal saber si hay luz u oscuridad y de dónde viene la luz, pero no pueden formar imágenes claras.
Ojos Complejos: Formando Imágenes
Existen ojos más complejos que sí pueden formar imágenes. Estos ojos tienen células que detectan la luz, células con pigmentos (color) y lentes. Se pueden dividir en dos clases principales:
- Ojos con invaginación cóncava: En su forma más sencilla, son como una pequeña copa con una abertura, como el ojo del Nautilus, un tipo de cefalópodo. Los más avanzados, como los de los vertebrados (incluidos nosotros), tienen lentes que ayudan a enfocar la luz. Otros, como los de las ostras, usan un espejo para reflejarla.
- Ojos compuestos: Estos se encuentran en insectos y crustáceos. Están formados por muchas unidades pequeñas llamadas omatidios, cada una con sus propias células que detectan la luz y, a veces, una lente.
Células que Captan la Luz: Fotorreceptores
Dentro de los ojos, hay células especiales llamadas fotorreceptoras que son las encargadas de captar la luz. Se distinguen dos grupos principales: los rabdómeros y los cuerpos ciliares. Se diferencian en varios aspectos:
- Usan diferentes "caminos" para procesar la señal de la luz.
- Los pigmentos que captan la luz (llamados opsinas) y otras proteínas son específicos para cada tipo de fotorreceptor.
- Responden de forma opuesta a la luz: un tipo de célula se "apaga" y el otro se "enciende" cuando recibe luz.
La Formación de las Lentes del Ojo
Las lentes son cruciales para enfocar la luz y formar imágenes claras.
Lentes en Vertebrados
En los vertebrados, las lentes están hechas de células especiales que contienen muchas proteínas llamadas cristalinas.
Lentes en Invertebrados
En los invertebrados, las lentes tienen una proteína que es producida por células especializadas del ojo.
La función de las lentes depende de cómo se distribuyen estas proteínas para crear un "gradiente" (un cambio gradual) en su capacidad de desviar la luz. Este gradiente es muy importante para la visión, especialmente en animales que viven en el agua. La forma en que se organizan las células para formar las lentes parece ser un ejemplo de "evolución convergente", lo que significa que diferentes especies desarrollaron soluciones similares de forma independiente.
El Gen Pax6: Un Director de Orquesta para el Ojo
El gen Pax6 es como un director de orquesta muy importante en el desarrollo del ojo. Juega un papel clave desde las primeras etapas de formación del ojo en el embrión hasta que el ojo está completamente desarrollado. Se activa en diferentes partes del ojo a medida que se forma, como la retina y la córnea.
El control genético de la formación de los ojos es complejo, como una red de genes que trabajan juntos. Sin embargo, Pax6 es tan importante que se le ha llamado un 'gen maestro'. Esto se debe a que, si se introduce el gen Pax6 de un animal en otro, como una mosca de la fruta, puede hacer que le crezcan ojos en lugares inesperados. Lo curioso es que, aunque el gen venga de otro animal, los ojos que se forman siempre tienen las características del ojo de la mosca. Esto sugiere que las diferencias en la forma de los ojos se deben más a cómo otros genes responden a Pax6 que al propio gen Pax6.
La Evolución del Ojo en Artrópodos
Los artrópodos (como insectos, arañas y crustáceos) tienen una gran variedad de ojos. Es sorprendente que los crustáceos tengan tantos tipos de ojos simples y compuestos, mientras que los arácnidos (excepto algunas arañas) solo tienen ojos simples. Los insectos suelen tener ojos compuestos, aunque muchos también tienen ojos simples llamados ocelos.
Aunque los ojos compuestos se consideran una característica antigua en los artrópodos, algunos miembros de este grupo los han perdido con el tiempo. Se pensaba que esto se debía a una reducción en los genes Pax6, pero estudios recientes muestran que no es así. La pérdida de ojos compuestos en algunos artrópodos se debe más a cambios en cómo se regulan otros genes que participan en la formación del ojo.
En algunos Diplopoda (milpiés), pueden tener ojos laterales adicionales dentro de la cabeza, que no están en contacto con la superficie. No se sabe con certeza cuál es su función o si son una característica antigua.
Estudiar cómo se forman los ojos en fósiles como los trilobites (animales marinos antiguos) nos ayuda a entender cómo evolucionaron los ojos en los animales actuales. Por ejemplo, en los trilobites, el área visual crecía a medida que el animal se desarrollaba, y las células migraban para formar las lentes.
El Origen de los Ojos en los Animales
Algunos científicos creen que los ojos pudieron haber evolucionado más de 65 veces de forma independiente en la historia de los animales. Por ejemplo, el ojo en cámara de los cefalópodos (como los pulpos) se parece mucho al ojo humano, pero no provienen de un mismo ancestro. Son un ejemplo de "homoplasia", donde estructuras similares evolucionan por separado.
Sin embargo, hay mucha evidencia de que genes como Pax6 y Six son muy parecidos en grupos tan diferentes como los artrópodos y los vertebrados. Esto sugiere que, a pesar de las grandes diferencias en la forma de sus ojos, podrían tener un origen común a partir de un ancestro que tenía ojos muy simples, solo para detectar la luz.
Estudiar organismos clave, como los Tunicados o los cnidarios (como las medusas), que se consideran grupos muy antiguos de animales, puede darnos muchas pistas sobre cómo se desarrollaron y evolucionaron los ojos. Por ejemplo, en las medusas, se cree que la retina apareció por primera vez.
El estudio de la evolución del ojo es un campo abierto y emocionante. La complejidad y perfección de los ojos fueron, en su momento, un argumento en contra de la teoría de la selección natural de Charles Darwin, pero hoy en día, la ciencia nos ayuda a entender cómo estas maravillosas estructuras pudieron surgir y diversificarse a lo largo de millones de años.
Galería de imágenes
-
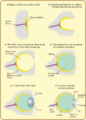
Principales etapas en la evolución del ojo en los vertebrados.

Véase también
 En inglés: Evolution of the eye Facts for Kids
En inglés: Evolution of the eye Facts for Kids
