
Proteína chaperona para niños
Las proteínas chaperonas son como "ayudantes" especiales dentro de nuestras células. Su trabajo principal es asegurarse de que otras proteínas, que se acaban de formar, se plieguen de la manera correcta. Imagina que una proteína es como una hoja de papel que necesita doblarse de una forma muy específica para funcionar bien; las chaperonas son las que la ayudan a doblarse perfectamente.
Estas proteínas chaperonas no se convierten en parte de la proteína que están ayudando. Solo se unen a ella por un tiempo para guiarla en su plegamiento, ayudarla a unirse con otras partes si es necesario, y a veces, a transportarla a su lugar de trabajo dentro de la célula. Los cambios en la forma tridimensional de una proteína pueden necesitar la ayuda de varias chaperonas trabajando juntas.
Las chaperonas son muy comunes y se encuentran en todas las células, tanto en las más simples (como las procariotas) como en las más complejas (como las eucariotas). Sus funciones principales son:
- Ayudar a las proteínas nuevas a plegarse correctamente.
- Ayudar a las proteínas que se han "desdoblado" (por ejemplo, por calor) a recuperar su forma.
- Colaborar en el plegamiento de proteínas que se mueven a diferentes partes de la célula.
Es importante saber que las chaperonas no le dicen a la proteína cómo plegarse. La información para que una proteína adquiera su forma correcta ya está en su propia estructura inicial. Las chaperonas solo se unen a las partes "pegajosas" de las proteínas mal plegadas para evitar que se peguen a otras proteínas o se doblen de forma incorrecta. También les dan estabilidad y tiempo para que encuentren su forma funcional.
Proteínas Chaperonas: Las Ayudantes de las Células
Las proteínas chaperonas son esenciales para la vida de las células. Sin ellas, muchas proteínas no podrían funcionar bien, lo que afectaría el buen funcionamiento de todo el organismo.
¿Qué Hacen las Proteínas Chaperonas?
Las chaperonas son como "entrenadores" de proteínas. Cuando una proteína se fabrica, es como una cadena larga. Para que funcione, debe doblarse en una forma tridimensional muy específica. Si no se dobla bien, no puede hacer su trabajo. Las chaperonas se aseguran de que este plegamiento sea perfecto.
Tipos Principales de Chaperonas
Existen diferentes familias de chaperonas, cada una con funciones específicas, pero todas con el objetivo de ayudar a otras proteínas.
Chaperoninas (Familia GroEL/GroES)
Las chaperoninas son un tipo de chaperonas muy interesantes. Un ejemplo es la GroEL en bacterias como Escherichia coli. Estas proteínas tienen una forma de cilindro hueco. Su trabajo es ayudar a plegar proteínas que han sido llevadas a lugares como los cloroplastos o las mitocondrias.
Se dividen en dos grupos principales:
- Las proteínas Hsp60 (como GroEL), que forman el cilindro hueco. Cada cilindro está hecho de dos anillos, y cada anillo tiene 7 piezas.
- Las proteínas Hsp10 (como GroES), que son como una "tapa" que se une a uno de los anillos de GroEL.
Hsp70: Las Rescatadoras de Proteínas
La familia Hsp70 (proteínas de choque térmico de 70 kilodaltons) es muy importante. Un ejemplo es la DnaK en E. coli. Se les llama "de choque térmico" porque se descubrieron por primera vez en células expuestas a altas temperaturas, donde aparecen rápidamente para ayudar a las proteínas dañadas.
Estas chaperonas trabajan junto con otras proteínas llamadas co-chaperonas, como Hsp40. Juntas, ayudan a:
- Deshacer el plegamiento incorrecto de proteínas.
- Evitar que las proteínas se peguen entre sí.
- Facilitar el plegamiento correcto de proteínas nuevas.
- Ayudar a las proteínas a moverse a través de las membranas de las mitocondrias y los cloroplastos.
Hsp90: Estabilizadoras Clave
Las chaperonas Hsp90 (como HtpG en E. coli) son también muy importantes para la supervivencia de las células. Representan una parte significativa de las proteínas en las células eucariotas.
Su función principal es ayudar a plegar, estabilizar y activar proteínas que son clave para la comunicación dentro de la célula. Esto incluye proteínas que transmiten señales, como las que controlan el crecimiento celular, y proteínas que forman la estructura de la célula.
Nucleoplasminas: Organizadoras del ADN
Las nucleoplasminas son proteínas que se encuentran en el núcleo de la célula. Su trabajo es ayudar a que el ADN se enrolle correctamente alrededor de unas proteínas llamadas histonas. Este enrollamiento es crucial para formar los nucleosomas, que son las unidades básicas de los cromosomas.
¿Cómo Funciona el Complejo GroEL/GroES?
El complejo GroEL/GroES es un ejemplo fascinante de cómo las chaperonas ayudan a las proteínas a plegarse. Así es como funciona:
- La proteína GroEL forma una estructura con forma de barril hueco. Uno de los anillos de GroEL se une a una molécula de energía llamada ATP y a la proteína que necesita plegarse.
- Luego, la "tapa" GroES se une a GroEL. Esto hace que la proteína que se va a plegar se libere dentro del barril. Dentro de este espacio, la proteína tiene un ambiente seguro para plegarse, aislada de otras proteínas que podrían interferir.
- La proteína tiene unos segundos para plegarse. Durante este tiempo, la energía del ATP se usa.
- Cuando se une una nueva proteína y más ATP al otro lado del barril, la proteína que ya se estaba plegando es liberada. Si no se plegó bien, el ciclo puede repetirse hasta que adquiera su forma correcta.

Galería de imágenes
-
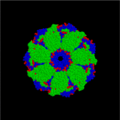
Vista cenital del complejo bacteriano de las chaperonas GroES/GroEL.
Véase también
 En inglés: Chaperone (protein) Facts for Kids
En inglés: Chaperone (protein) Facts for Kids
