
Plasticidad neuronal para niños
La plasticidad neuronal, también conocida como neuroplasticidad, es la increíble habilidad de tu cerebro para cambiar y adaptarse a nuevas experiencias y aprendizajes. Imagina que tu cerebro es como una red de caminos que se pueden modificar: la plasticidad neuronal permite que se creen nuevos caminos o que los existentes se hagan más fuertes o más débiles. Esto sucede porque las neuronas, que son las células de tu cerebro, pueden formar nuevas conexiones entre sí o ajustar las que ya tienen. Gracias a esto, tu cerebro puede aprender cosas nuevas, recordar información y adaptarse a lo que sucede a tu alrededor.


Contenido
¿Cómo se comunican las neuronas?
Las neuronas se comunican entre sí en lugares especiales llamados sinapsis. Piensa en una sinapsis como un pequeño puente donde una neurona envía un mensaje a otra. Este mensaje se transmite de forma eléctrica y química.
Cuando una señal eléctrica llega al final de una neurona (la presináptica), esta libera unas sustancias químicas llamadas neurotransmisores en el espacio entre las dos neuronas. Estos neurotransmisores son como llaves que encajan en cerraduras especiales (llamadas receptores) en la siguiente neurona (la postsináptica).
Tipos de acción en la sinapsis
Dependiendo del neurotransmisor y del receptor, la neurona que recibe el mensaje puede reaccionar de diferentes maneras:
Acción Ionotrópica: Mensajes rápidos
Algunos neurotransmisores abren directamente "canales" en la membrana de la neurona receptora. Estos canales permiten que pequeñas partículas cargadas (iones) entren o salgan de la célula, cambiando su carga eléctrica.
- Potencial Excitador Postsináptico (PEPS): Si entran iones positivos, la neurona se "excita" y es más probable que envíe su propio mensaje eléctrico. Por ejemplo, el glutamato es un neurotransmisor que suele causar este efecto.
- Potencial Inhibidor Postsináptico (PIPS): Si entran iones negativos, la neurona se "inhibe" y es menos probable que envíe un mensaje. El GABA es un ejemplo de neurotransmisor que "apaga" la neurona.
Acción Metabotrópica: Mensajes con efectos duraderos
Otros neurotransmisores no abren canales directamente, sino que activan un proceso dentro de la neurona que puede tener efectos más duraderos. Es como si la llave no abriera una puerta, sino que encendiera una máquina dentro de la casa. Esto puede hacer que los canales se abran por más tiempo o que se muevan a lugares donde la comunicación sea más efectiva.
Neurotransmisión primaria y secundaria
- La neurotransmisión primaria es la acción rápida, como la que ocurre con los receptores ionotrópicos, que hacen que una neurona se excite o se inhiba rápidamente.
- La neurotransmisión secundaria es la acción más lenta y duradera, como la de los receptores metabotrópicos, que pueden cambiar la fuerza o la probabilidad de que un mensaje se transmita con éxito.
La combinación de estas dos formas de comunicación es lo que permite la plasticidad neuronal.
¿Cómo integra la información el cerebro?
Tu cerebro recibe muchísima información al mismo tiempo. Una sola neurona puede recibir miles de conexiones de otras neuronas. Para entender todo esto, las neuronas "suman" todas las señales que reciben, tanto las que las excitan como las que las inhiben.
- Suma espacial: Imagina que muchas neuronas envían mensajes a una sola neurona al mismo tiempo. La neurona receptora suma todos esos mensajes para decidir si debe enviar su propia señal.
- Suma temporal: Si una neurona envía muchos mensajes muy seguidos a otra, la neurona receptora también los suma a lo largo del tiempo para decidir su respuesta.
La forma en que las neuronas se conectan no está totalmente predeterminada. Tus experiencias pueden cambiar cómo se forman y fortalecen estas conexiones.


Plasticidad y el aprendizaje
La plasticidad neuronal es clave para el aprendizaje y la memoria. Cuando aprendes algo nuevo, las conexiones entre tus neuronas cambian.
- Facilitación sináptica: Si una neurona envía mensajes repetidamente a otra, la conexión entre ellas puede volverse más fuerte, haciendo que el siguiente mensaje sea más efectivo.
- Depresión sináptica: Por el contrario, si los mensajes se repiten mucho y de cierta manera, la conexión puede debilitarse.
Estas modificaciones en la fuerza de las conexiones pueden durar desde minutos hasta días, semanas o incluso años, lo que nos permite formar recuerdos a largo plazo.
Ejemplos de plasticidad en animales
Los científicos han estudiado la plasticidad en animales más simples para entender cómo funciona. Un ejemplo famoso es el caracol marino Aplysia.
Habituación y sensibilización
- Habituación: Si tocas suavemente el sifón de la Aplysia muchas veces, el caracol se acostumbra y deja de retraer su branquia. Esto ocurre porque la conexión entre la neurona que detecta el toque y la neurona que mueve la branquia se debilita.
- Sensibilización: Si después de tocar el sifón, le das un pequeño golpe en la cola, el caracol se vuelve más sensible y vuelve a retraer la branquia con fuerza al tocar el sifón. Esto sucede porque se activan otras neuronas que liberan sustancias que fortalecen temporalmente la conexión original.
Estos cambios, tanto la habituación como la sensibilización, pueden durar mucho tiempo, lo que demuestra una forma simple de memoria.



Plasticidad a largo plazo en el cerebro
En el cerebro de los mamíferos, incluyendo el humano, existen formas de plasticidad que duran mucho tiempo y son fundamentales para la memoria.
Potenciación a largo plazo (PLP)
La PLP es un aumento duradero en la fuerza de las conexiones sinápticas. Se ha estudiado mucho en una parte del cerebro llamada hipocampo, que es muy importante para formar nuevos recuerdos. Si ciertas neuronas en el hipocampo se estimulan de una manera específica y rápida, sus conexiones se vuelven mucho más fuertes por un largo tiempo. Esto ocurre gracias a la entrada de calcio en las neuronas, que activa proteínas que fortalecen la sinapsis.
Depresión a largo plazo (DLP)
La DLP es lo contrario de la PLP: una disminución duradera en la fuerza de las conexiones sinápticas. También ocurre en el hipocampo y en el cerebelo, una parte del cerebro importante para el aprendizaje de movimientos. La DLP es necesaria para que el cerebro pueda "borrar" o debilitar conexiones que ya no son útiles, permitiendo un aprendizaje más flexible.
Tanto la PLP como la DLP son cruciales para que el cerebro pueda aprender y recordar. Los estudios han demostrado que las moléculas y los procesos involucrados en la PLP y la DLP también son esenciales para el aprendizaje y la memoria en general.
El cerebro adulto también cambia
Durante mucho tiempo se pensó que el cerebro adulto no cambiaba mucho, pero ahora sabemos que es muy dinámico y adaptable. Incluso en la edad adulta, tu cerebro sigue siendo capaz de reorganizarse y formar nuevas conexiones, lo que te permite seguir aprendiendo y adaptándote a lo largo de toda tu vida.
Galería de imágenes
-
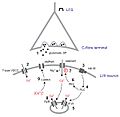
Interacción neurotranmisora.
-

A. Vista esquemática de un potencial de acción ideal, mostrando sus distintas fases. B. Registro real de un potencial de acción, normalmente deformado, comparado con el esquema debido a las técnicas electrofisiológicas utilizadas en la medición.


Véase también
 En inglés: Neuroplasticity Facts for Kids
En inglés: Neuroplasticity Facts for Kids
- Neuroepigenética
- Eric Kandel
- Transducción de señales
- Receptor de glutamato
- Neurodegeneración
