
Osmorregulación para niños
La Osmorregulación es un proceso vital que usan los seres vivos para mantener el equilibrio de agua y sales dentro de su cuerpo. Imagina que tu cuerpo es como una piscina con una cantidad perfecta de agua y sal; la osmorregulación se encarga de que esa cantidad se mantenga siempre así.
Todas las reacciones químicas que nos mantienen vivos necesitan un ambiente líquido muy específico. Por eso, es súper importante que la cantidad de agua y de sustancias disueltas (llamadas solutos) se mantenga dentro de límites muy precisos. La osmorregulación es clave para lograr este equilibrio, controlando cómo se mueven los solutos y el agua entre el interior del cuerpo y el ambiente exterior.
Este proceso se basa en la ósmosis, que es el movimiento del agua a través de una membrana especial (llamada semipermeable). El agua siempre se mueve desde donde hay más agua (y menos solutos) hacia donde hay menos agua (y más solutos), hasta que las concentraciones se igualan. La cantidad de solutos en una solución se mide en miliosmoles por litro (mOsm).
Contenido
Osmorregulación en Células
El control del agua es diferente en las células de los animales y en las de las plantas. Aunque la difusión y la ósmosis son importantes en ambos, hay otros factores como la presión osmótica.
Células Animales
En la mayoría de los animales, los líquidos que rodean las células tienen una concentración de solutos similar a la de los líquidos dentro de las células. Esto es importante porque si la concentración fuera muy diferente, la célula podría hincharse y explotar, o arrugarse y encogerse. Para evitar esto, las células animales usan energía para bombear constantemente ciertas sales, como el sodio, hacia afuera.
Células Vegetales
Las células vegetales tienen una pared celular rígida que les permite acumular más solutos dentro que fuera. Esto hace que el agua tienda a entrar en la célula por ósmosis, aumentando su volumen y una presión interna llamada presión de turgencia. La pared celular evita que la célula explote, ejerciendo una presión que equilibra el agua.
Las células vegetales y los hongos usan un sistema diferente al de los animales para mover sustancias. En lugar de bombas de sodio y potasio, las plantas usan bombas de protones (iones de hidrógeno) para crear un equilibrio que les permite mover otros solutos.
Osmorregulación en Animales
La osmorregulación en animales depende mucho de unas células especiales llamadas epitelios de transporte.
Epitelios de Transporte
Estos epitelios son como "puertas" especializadas que regulan el movimiento de los solutos. Su función principal es crear diferencias de carga eléctrica que permiten transportar iones y otras sustancias en una dirección específica. Esto lo logran con la ayuda de proteínas especiales (canales, bombas) que están distribuidas de forma desigual en sus membranas. Este transporte consume energía.
Osmorregulación en Diferentes Ambientes
Los animales se han adaptado a vivir en muchos tipos de ambientes acuáticos, desde agua dulce (con pocos solutos) hasta agua de mar (con mucha sal). Por eso, los desafíos para regular el agua son muy distintos.
Algunos animales solo toleran un rango pequeño de salinidad (estenohalinos), mientras que otros pueden vivir en un rango muy amplio (eurihalinos).
Los animales acuáticos, como los invertebrados y los peces, usan órganos como las branquias, los riñones y, en algunos casos, el intestino o glándulas especiales para regular el agua y las sales.
Existen dos estrategias principales:
- Osmoconformistas: Estos animales tienen una concentración de solutos similar a la del agua donde viven. Sus cuerpos son permeables al agua y a las sales, por lo que sus condiciones internas cambian con el ambiente. Suelen vivir en ambientes con salinidad muy estable.
- Osmorreguladores: Estos animales mantienen una concentración interna de solutos bastante estable, sin importar los cambios en el ambiente externo. Si tienen más solutos que el exterior (como los animales de agua dulce), se les llama hiperosmóticos. Si tienen menos solutos que el exterior (como los peces marinos), son hipoosmóticos.
La osmorregulación requiere energía. En muchos peces, puede consumir hasta el 5% de su energía en reposo.
Ambientes Acuáticos
Animales Marinos
La mayoría de los invertebrados marinos son osmoconformistas. Sin embargo, los vertebrados marinos (como los peces) suelen mantener una concentración de solutos en su sangre que es aproximadamente un tercio de la del agua de mar.
Hay tres grupos de vertebrados marinos según su osmorregulación:
- Isosmóticos con el agua de mar: Solo un grupo de peces sin mandíbula, los mixinoideos, tienen la sangre muy parecida al agua de mar. Otros, como los tiburones, son casi isosmóticos gracias a moléculas orgánicas como la urea. Estos peces ganan agua por ósmosis y eliminan el exceso de sales por los riñones y una glándula rectal.
- Hipoosmóticos: Los peces óseos marinos (teleósteos) tienen fluidos corporales más diluidos que el agua de mar. Pierden agua por ósmosis y ganan sales por difusión. Para contrarrestarlo:
* Beben agua salada. * Absorben agua en su tubo digestivo. * Eliminan el exceso de sales activamente a través de sus branquias, usando células especiales llamadas células de cloruro. * Excretan una orina muy concentrada en sales divalentes, pero en muy poca cantidad.
- Tetrápodos marinos: Aves y reptiles marinos no pueden producir una orina muy concentrada, por lo que tienen glándulas de la sal que excretan grandes cantidades de sal. Los mamíferos marinos, como las ballenas, sí pueden producir una orina más concentrada que el agua de mar, lo que les permite obtener agua neta al beberla.
Animales de Agua Dulce
Los animales de agua dulce tienen fluidos corporales más concentrados que su entorno. Esto significa que el agua tiende a entrar en su cuerpo y la sal tiende a salir. Para manejar esto, son reguladores hiperosmóticos y usan los siguientes mecanismos:
- Poca permeabilidad: Su piel o superficie es menos permeable al agua y a las sales.
- Orina diluida: Excretan grandes cantidades de orina muy diluida para eliminar el exceso de agua.
- Captación de iones: Tienen mecanismos para absorber sales del agua, especialmente a través de las branquias en los peces.
Reguladores Hiperosmóticos-Hipoosmóticos
Algunos peces, como los que viven en estuarios o migran entre agua dulce y salada, pueden cambiar su forma de osmorregular. Regulan de una manera en agua dulce y de otra en agua salada. Este cambio está controlado por hormonas.
Ambientes Terrestres
El mayor desafío para los animales terrestres es no deshidratarse. Su concentración de solutos es intermedia entre la de los animales marinos y los de agua dulce.
Los animales terrestres obtienen agua de tres maneras: bebiéndola, comiéndola (en los alimentos) y produciéndola en su metabolismo. Pierden agua al orinar, defecar, por evaporación al respirar y a través de la piel. El equilibrio de agua depende de la humedad, temperatura y viento.
Se dividen en dos grupos:
- Humídicos: Se deshidratan rápidamente en ambientes secos y necesitan vivir en lugares húmedos. Ejemplos: lombrices de tierra, babosas, anfibios.
- Xéricos: Pueden vivir en climas secos con poca agua. Ejemplos: reptiles, aves, mamíferos, insectos, arácnidos.
Algunos animales, como los camellos, son excepcionales y pueden tolerar una gran pérdida de agua corporal.
Mecanismos Adaptativos de Conservación de Agua
- Comportamiento: Buscan lugares húmedos, son activos de noche o al atardecer, y eligen alimentos con más agua.
- Especializaciones físicas: Tienen pieles o cubiertas poco permeables al agua (como la cutícula de los insectos o la piel queratinizada de los reptiles). Algunos mamíferos del desierto tienen mecanismos en sus vías respiratorias para recuperar el vapor de agua al exhalar. Los insectos tienen válvulas en sus espiráculos para evitar la pérdida de agua.
- Reducción de pérdidas fecales: Extraen agua de las heces antes de eliminarlas.
- Latencia: Algunos animales entran en un estado de inactividad cuando hay escasez de agua, reduciendo su metabolismo y, por lo tanto, la pérdida de agua y la producción de desechos.
La eliminación de desechos nitrogenados (como el amoníaco, que es tóxico) es un problema para la conservación del agua.
- Los animales acuáticos suelen excretar amoníaco directamente en el agua (amoniotélicos).
- Los animales terrestres lo transforman en urea (ureotélicos, como mamíferos y anfibios) o en ácido úrico (uricotélicos, como insectos, reptiles y aves). El ácido úrico requiere muy poca agua para ser eliminado.
- Obtención de agua del aire: Unos pocos artrópodos pueden extraer agua del aire no saturado, incluso en ambientes muy secos.
Órganos Reguladores
En los animales terrestres, los órganos excretores no solo eliminan desechos, sino que también son clave en la osmorregulación. Regulan el contenido de agua y sales. Incluyen nefridios (en platelmintos y anélidos), tubos de Malpighi (en insectos y arañas) y los riñones (en vertebrados).
Todos estos sistemas comparten principios básicos:
- Ultrafiltración: La presión de la sangre fuerza el paso de líquidos a través de un filtro que retiene células y moléculas grandes, dejando pasar agua, iones y solutos pequeños.
- Secreción activa: En algunos animales (como insectos), los iones son bombeados activamente al órgano excretor, arrastrando agua y otros solutos.
Animales humídicos
- Platelmintos: Tienen protonefridios con células flamígeras que mueven el fluido hacia afuera. Reabsorben sales para mantener el equilibrio.
- Anélidos: Tienen metanefridios que filtran el líquido del cuerpo y reabsorben sales.
- Anfibios: Sus riñones funcionan como los de los peces de agua dulce, excretando orina diluida. En tierra, reabsorben agua a través de la piel de la vejiga.
Animales xéricos Producen orina más concentrada que su plasma sanguíneo.
- Insectos y arañas: Usan los tubos de Malpighi y glándulas rectales. Las sales son secretadas activamente en los tubos, arrastrando agua y desechos. Luego, la mayor parte del agua y el potasio se reabsorben en el recto.
- Vertebrados: El riñón es el órgano principal, y su unidad funcional es el nefrón.
* Nefrón: Tiene un componente vascular (el glomérulo, que filtra la sangre) y un componente tubular (donde el filtrado se convierte en orina). * Filtración: La sangre se filtra en el glomérulo, dejando pasar agua y solutos pequeños. * Secreción: Sustancias se mueven de la sangre a los túbulos de forma selectiva. * Reabsorción: La mayoría del agua y los solutos útiles (como glucosa) se reabsorben de los túbulos de vuelta a la sangre.
Los riñones de mamíferos y aves tienen nefrones especiales (yuxtamedulares) con asas de Henle largas que son clave para concentrar la orina y ahorrar agua. Cuanto más larga es el asa de Henle, mayor es la capacidad de concentrar la orina.
Control del Equilibrio Hídrico en Animales Terrestres
Los riñones y otros órganos están controlados por hormonas, que a su vez son reguladas por sistemas de retroalimentación.
- Hormona Antidiurética (ADH): Producida en el cerebro, esta hormona aumenta la permeabilidad al agua de los túbulos renales, haciendo que se reabsorba más agua y se produzca menos orina concentrada.
- Sistema Renina-Angiotensina-Aldosterona (SRAA): Este sistema se activa cuando disminuye la sal, el volumen de sangre o la presión sanguínea. La renina (una enzima del riñón) inicia una cadena de reacciones que lleva a la producción de angiotensina II (que aumenta la presión arterial) y aldosterona (que aumenta la reabsorción de sodio y, por lo tanto, de agua).
- Péptido Natriurético Atrial (PNA): Producido por el corazón cuando la presión es alta, esta hormona dilata las arterias renales, inhibe la aldosterona y la ADH, y promueve la pérdida de sodio y agua en la orina, reduciendo la presión sanguínea.
Mecanismos de Aumento de Presión
Cuando el cuerpo necesita aumentar la presión sanguínea:
- El hipotálamo (en el cerebro) produce la hormona ADH, que se libera desde la hipófisis. La ADH aumenta la permeabilidad de los riñones al agua y causa que las arterias se contraigan, subiendo la presión.
- La hipófisis también produce la hormona ACTH, que estimula las glándulas suprarrenales para liberar aldosterona. La aldosterona hace que el riñón retenga sodio, que pasa a la sangre, aumentando la presión.
- El riñón produce la enzima renina, que ayuda a formar angiotensina II, manteniendo el ciclo de aumento de presión.
Mecanismos de Disminución de Presión
El corazón es el principal órgano que disminuye la presión arterial. Cuando la presión es demasiado alta, el corazón produce el péptido natriurético auricular, que inhibe la producción de aldosterona, lo que lleva a una menor retención de sodio y agua, y por lo tanto, a una disminución de la presión.
Osmorregulación en Plantas
Las plantas se clasifican según sus necesidades de agua:
- Hidrófitas: Viven en el agua.
- Mesófitas: Necesitan una cantidad moderada de agua.
- Xerófitas: Adaptadas a ambientes secos.
- Halófitas: Toleran altas concentraciones de sal en el suelo.
Las plantas xerófitas y halófitas están constantemente bajo estrés hídrico. Se adaptan con estrategias como el uso eficiente del agua, cutículas gruesas, estomas escondidos y alta densidad de pelos. Las halófitas mantienen un bajo potencial osmótico en sus células para absorber agua y evitan el daño por sal almacenando iones en vacuolas o excretándolos.
Las plantas responden al estrés hídrico a nivel celular y molecular, aumentando la producción de osmolitos (sustancias que ayudan a equilibrar el agua), acuaporinas (canales de agua) y ácido abscísico (una hormona).
Acumulación de Osmolitos como Respuesta Osmorreguladora
Para resistir la falta de agua, las plantas acumulan solutos especiales en sus células. Esto se llama ajuste osmótico y es clave para que las plantas se adapten a la sequía o la salinidad. Este ajuste permite a las plantas absorber agua de suelos más secos y mantener su presión interna. Los solutos compatibles u osmolitos son moléculas orgánicas que no dañan las funciones de la célula, como la prolina o el sorbitol.
Regulación de la Permeabilidad Celular
Las acuaporinas son proteínas que forman canales de agua en las membranas de las células vegetales. Son fundamentales para el movimiento del agua desde las raíces hasta las hojas. Su cantidad y actividad pueden cambiar según las condiciones ambientales para ayudar a la planta a obtener o retener agua.
Control Estomático de la Pérdida de Agua
Las plantas primitivas, como líquenes y musgos, no tienen mecanismos para evitar la pérdida de agua y solo son activas cuando hay humedad. Las plantas vasculares tienen una cutícula impermeable en sus hojas, pero la mayor parte del agua se pierde a través de los estomas.
Los estomas son pequeños poros en las hojas, rodeados por dos células especiales llamadas células guarda. Estas células controlan la apertura y el cierre del poro. Cuando los estomas están abiertos, el agua se evapora y el dióxido de carbono entra a la hoja. Durante el estrés hídrico, los estomas se cierran para reducir la pérdida de agua.
Hay dos mecanismos de cierre:
- Mecanismo hidropasivo: Se debe a la pérdida directa de agua por evaporación de las células guarda, lo que las hace perder turgencia y cerrar el poro.
- Mecanismo hidroactivo: Los estomas se cierran en respuesta a señales como el estrés hídrico. La hormona ácido abscísico (ABA) juega un papel clave, activando procesos que reducen el contenido de solutos en las células guarda, haciendo que pierdan agua y se cierren.
Respuesta de las Plantas Halófitas al Estrés Salino
La mayoría de las plantas no toleran altas concentraciones de sal, pero las halófitas sí. El estrés salino afecta a las plantas de dos maneras:
- Osmótico: Si el suelo tiene mucha sal, la planta no puede absorber agua. Las halófitas responden con ajuste osmótico.
- Iónico: El sodio y el cloro son tóxicos en altas concentraciones.
Las halófitas pueden ser:
- Exclusivas de iones: Limitan la cantidad de sal que llega a sus hojas.
- Inclusivas de iones: Absorben mucha sal y la almacenan en vacuolas de células especiales, o la excretan a la superficie de las hojas a través de glándulas de sal.
Galería de imágenes
-

Movimiento de agua y de iones en un pez teleósteo marino
-
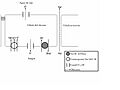
Modelo de la expulsión de ClNa por las células de cloruro de las branquias de peces teleósteos
-

Movimiento de agua y de iones en pez teleósteo de agua dulce
-
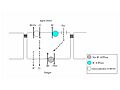
Modelo de la captación de ClNa por el epitelio branquial de peces teleósteos
-

Balance hídrico en un animal terrestre
-

Mecanismo de transporte de sodio, potasio y cloro en la rama gruesa ascendente del asa de Henle.
-

Control por retroalimentación negativa de la secreción de la hormona antidiurética (ADH)
-

Modelo del transporte celular en la reabsorción de sodio en una célula P sensible a aldosterona.
-

Modelo de los procesos de transporte de membrana durante el cierre de los estomas mediado por ABA. 1. El ABA se une al receptor en la superficie de la membrana plasmática de las células guarda.2.El ABA activa la apertura de canales Ca2+ y aumenta el Ca2+ citosólico. 3.El aumento del Ca2+ citosólico inhibe a las bombas protónicas. 4.El aumento del Ca2+ citosólico inhibe a los canales K+ de entrada. 5.El aumento del Ca2+ citosólico activa dos tipos de canales aniónicos de membrana y se despolariza la membrana plasmática. 6.La despolarización activa canales K+ de salida.






.png)


Véase también
 En inglés: Osmoregulation Facts for Kids
En inglés: Osmoregulation Facts for Kids
