
Microcompartimiento bacteriano para niños
Los microcompartimientos bacterianos (también conocidos como MCB o BMC por sus siglas en inglés) son como pequeñas "cajas" dentro de las bacterias. Están hechos de proteínas y guardan en su interior otras proteínas y enzimas importantes. Estos compartimientos suelen medir entre 40 y 200 nanómetros de diámetro, ¡son muy, muy pequeños! Su cubierta funciona de manera parecida a una membrana, controlando qué sustancias pueden entrar y salir.
Existen otros compartimientos similares en bacterias y arqueas, como los nanocompartimientos de encapsulina y las vesículas de gas. Incluso los eucariotas (organismos con células más complejas, como las plantas y los animales) tienen estructuras parecidas, como el complejo de bóveda.
Aunque la cubierta de los microcompartimientos bacterianos se parece a las cápsides de los virus (las cubiertas que protegen el material genético de los virus), no están relacionados. Los estudios sugieren que los virus podrían haber surgido de estructuras parecidas a estos microcompartimientos hace mucho tiempo, antes de que existieran las células tal como las conocemos.
Contenido
- ¿Cómo se descubrieron los microcompartimientos bacterianos?
- ¿De qué están hechas las cubiertas de los microcompartimientos?
- ¿Cómo funciona la permeabilidad de la cubierta?
- Tipos de microcompartimientos bacterianos
- ¿Cómo se forman los microcompartimientos?
- Regulación de los microcompartimientos
- Aplicaciones en biotecnología
- Galería de imágenes
- Véase también
¿Cómo se descubrieron los microcompartimientos bacterianos?
Los primeros microcompartimientos bacterianos se vieron en la década de 1950 usando microscopios electrónicos en cianobacterias. Al principio, se les llamó carboxisomas cuando se descubrió que ayudaban a las bacterias a fijar el carbono (un proceso importante para su alimentación).
Hasta los años 90, se pensaba que los carboxisomas eran algo raro, solo presente en algunas bacterias que fabrican su propio alimento. Pero luego, se encontraron genes similares a los de la cubierta de los carboxisomas en bacterias que usan propanodiol o etanolamina. Más tarde, se vieron estructuras parecidas a los carboxisomas en células de Salmonella que crecían con propanodiol. A estos nuevos compartimientos se les llamó metabolosomas, porque están relacionados con el metabolismo (los procesos químicos de la célula).
Aunque los carboxisomas y los metabolosomas guardan enzimas diferentes y tienen funciones distintas, los genes que forman sus cubiertas son muy parecidos. La mayoría de los genes que codifican las proteínas de la cubierta y las enzimas que guardan se encuentran juntos en el ADN de las bacterias. Hoy en día, con la ayuda de la bioinformática (el uso de computadoras para estudiar información biológica), se han identificado más de 20.000 genomas bacterianos y se pueden encontrar todos los genes de estos compartimientos. En 2014, se identificaron 23 tipos diferentes de microcompartimientos con hasta 10 funciones distintas en 23 grupos de bacterias.
¿De qué están hechas las cubiertas de los microcompartimientos?
La cubierta de los microcompartimientos bacterianos tiene una forma de poliedro, como un balón de fútbol, y está hecha de varias piezas de proteínas. Las piezas principales son proteínas que se unen para formar hexágonos (seis lados) y pentágonos (cinco lados).
Proteínas hexagonales (BMC-H)
Las proteínas BMC-H son las más comunes en la cubierta. Se unen formando hexágonos con un pequeño agujero en el centro. Se cree que estos agujeros son como "puertas" que permiten el paso selectivo de moléculas pequeñas, llamadas metabolitos, a través de la cubierta.
Proteínas en tándem (BMC-T)
Otro tipo de proteínas de la cubierta, las BMC-T, forman trímeros (grupos de tres) que también tienen forma de hexágono. Algunas de estas proteínas pueden apilarse y tener poros que se abren y cierran, como una esclusa, para controlar lo que entra y sale. Algunas BMC-T también tienen grupos de hierro y azufre y podrían ayudar a transportar electrones a través de la cubierta.
Proteínas pentagonales (BMC-P)
Para formar una cubierta completa, se necesitan doce unidades pentagonales que se colocan en los vértices (las esquinas) de la estructura. Las proteínas de la familia EutN/CcmL suelen formar pentámeros (grupos de cinco) y son importantes para que la cubierta se forme correctamente. Por ejemplo, sin estas proteínas, algunos microcompartimientos no pueden formarse o se vuelven "con fugas".
¿Cómo funciona la permeabilidad de la cubierta?
Las enzimas están dentro de la cubierta de los microcompartimientos, y algunas sustancias deben entrar y salir para que funcionen. Por ejemplo, en los carboxisomas, el bicarbonato debe cruzar la cubierta, mientras que la difusión de dióxido de carbono y oxígeno está limitada. En los metabolosomas, el propionaldehído (una sustancia que puede ser dañina) se mantiene dentro para proteger la célula.
Se cree que los pequeños agujeros en las proteínas hexagonales de la cubierta son los canales por donde pasan los metabolitos. Por ejemplo, los poros de la cubierta del carboxisoma tienen una carga positiva que atrae al bicarbonato, que tiene carga negativa. En el microcompartimiento de PDU, se ha demostrado que el poro de una proteína específica es la vía de entrada para el propanodiol. Para metabolitos más grandes, algunas proteínas BMC-T tienen un mecanismo que las activa para permitir el paso.
Tipos de microcompartimientos bacterianos
Un estudio reciente encontró hasta diez funciones metabólicas diferentes que los microcompartimientos pueden realizar. La mayoría están involucradas en la fijación de carbono (carboxisomas) o en la oxidación de aldehídos (metabolosomas).
Carboxisomas: para fijar el carbono
Los carboxisomas guardan enzimas como la RuBisCO y la anhidrasa carbónica. Ayudan a las bacterias que fijan carbono a concentrar el dióxido de carbono. El bicarbonato entra en el carboxisoma y se convierte en dióxido de carbono, que es lo que usa la enzima RuBisCO. La cubierta del carboxisoma no deja salir fácilmente el dióxido de carbono, lo que aumenta su concentración y mejora la fijación de carbono.
Metabolosomas: para procesar aldehídos
Además de los carboxisomas, existen varios metabolosomas que ayudan a las bacterias a procesar aldehídos. Se cree que estos compartimientos guardan los aldehídos, que pueden ser tóxicos para las células. Los metabolosomas suelen tener tres enzimas principales: aldehído deshidrogenasa, alcohol deshidrogenasa y fosfotransacilasa. Estas enzimas ayudan a reciclar otras moléculas necesarias y a transformar los aldehídos en compuestos que la célula puede usar para crecer.
Microcompartimientos de PDU
Algunas bacterias usan 1,2-propanodiol como alimento. Para ello, utilizan un microcompartimiento que guarda varias enzimas de esta vía. El propionaldehído, un producto intermedio, es tóxico para las células, por lo que se guarda dentro del microcompartimiento. Los productos finales son propanol y propionato, que la bacteria puede usar para crecer.
Microcompartimientos con enzimas de radicales glicilo (GRM)
Se han encontrado microcompartimientos que contienen enzimas de radicales glicilo. Se ha demostrado que uno de estos tipos está involucrado en la fermentación de azúcares como la fucosa y la ramnosa, que se transforman en 1,2-propanodiol.
Microcompartimientos de función desconocida (BUF)
Algunos microcompartimientos no contienen las enzimas típicas de los carboxisomas o metabolosomas. Se cree que podrían estar involucrados en el procesamiento de compuestos que contienen nitrógeno.
¿Cómo se forman los microcompartimientos?
Carboxisomas
La formación de los beta-carboxisomas comienza con una proteína llamada CcmM, que agrupa a la enzima RuBisCO. Luego, otras proteínas se unen para formar la cubierta hexagonal. Los carboxisomas se organizan dentro de la célula para asegurarse de que se distribuyan por igual cuando la célula se divide.
La formación de los alfa-carboxisomas puede ser diferente. Se ha visto que pueden formarse a partir de la unión simultánea de enzimas y proteínas de la cubierta.
Metabolosomas
La formación de los metabolosomas es similar a la de los beta-carboxisomas. Las proteínas que se van a guardar se agrupan primero. Muchas de estas proteínas tienen "extensiones" que son parecidas a un péptido (una pequeña cadena de proteínas) que ayuda a reclutar las proteínas de la cubierta. Estos péptidos son cortos y pueden ayudar a que las enzimas se guarden dentro del microcompartimiento.
Regulación de los microcompartimientos
La mayoría de los microcompartimientos se activan solo cuando su "alimento" específico está presente. Por ejemplo, los microcompartimientos de PDU se activan cuando hay propanodiol o glicerol. Esto está controlado por proteínas reguladoras que detectan estas sustancias.
Los microcompartimientos de EUT se activan cuando hay etanolamina y vitamina B12 al mismo tiempo. Otros microcompartimientos se activan con la presencia de azúcares como la fucosa o la ramnosa.
Aplicaciones en biotecnología
Los microcompartimientos bacterianos son muy interesantes para la biotecnología (el uso de organismos vivos para crear productos o procesos útiles).
Como los carboxisomas mejoran la fijación de carbono, se está investigando cómo introducirlos en los cloroplastos de las plantas para que estas puedan capturar más dióxido de carbono, lo que podría ayudar a mejorar los cultivos.
Además, como las proteínas de la cubierta de los microcompartimientos se autoensamblan, se pueden crear "cajas vacías" y luego diseñarlas para que contengan otras sustancias. Se ha descubierto que un pequeño péptido puede ayudar a "encapsular" proteínas dentro de estos compartimientos. Por ejemplo, al añadir este péptido a ciertas enzimas, los científicos han logrado crear un "biorreactor" que produce etanol. También se pueden modificar los poros de la cubierta para controlar qué sustancias entran y salen.


Galería de imágenes
-
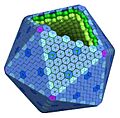
Ejemplo de un microcompartimiento bacteriano.
-
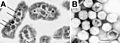
Microcompartimientos bacterianos a vista micróscopica.
Véase también
 En inglés: Bacterial microcompartment Facts for Kids
En inglés: Bacterial microcompartment Facts for Kids
