
Cucurbita maxima subsp. andreana para niños
El zapallito amargo o zapallo amargo (cuyo nombre científico es Cucurbita maxima subsp. andreana) es una planta silvestre que se encuentra en Argentina y Uruguay. Es el pariente salvaje de los zapallos y zapallitos que comemos, como el Zapallito redondo del tronco.
Esta planta produce un fruto pequeño y liviano, con una cáscara dura y una pulpa seca y fibrosa donde están las semillas. A diferencia de los zapallos cultivados, el zapallito amargo tiene un sabor muy amargo y puede ser perjudicial si se consume. Es la única calabacita silvestre que crece en Argentina y Uruguay.
Aunque es silvestre, puede cruzarse con los zapallos cultivados de la misma especie. Esto es importante porque, si el polen del zapallito amargo llega a las flores de los zapallos dulces, las semillas resultantes podrían producir plantas con frutos amargos en la siguiente cosecha. Por eso, a veces se le considera una "maleza" en los campos de cultivo.
Contenido
¿Dónde crece el zapallito amargo?
El zapallito amargo se encuentra principalmente en las regiones templadas de Argentina y Uruguay, donde los veranos son cálidos y los inviernos fríos. Se ha registrado su presencia en varias provincias argentinas como Buenos Aires, Entre Ríos, Santa Fe, Córdoba, La Pampa, San Luis, Santiago del Estero, La Rioja y Mendoza.
Esta planta prefiere crecer en lugares donde el suelo ha sido modificado, como terraplenes, maizales, antiguas casas abandonadas o cerca de madrigueras de animales como las vizcachas.
¿Cómo es el zapallito amargo?
El zapallito amargo es una planta anual, lo que significa que vive solo una temporada. Germina en primavera, produce flores y frutos durante el verano, y muere con las primeras heladas del otoño, dejando sus semillas para la próxima temporada.
Es una planta trepadora o rastrera, con tallos que se extienden y echan raíces en los nudos que tocan la tierra. Sus hojas son redondas, con forma de riñón o con tres lóbulos, y sus zarcillos (pequeños "brazos" que usa para sujetarse) se dividen en varias ramas. Algunas plantas tienen hojas verdes y otras tienen hojas con manchas.
-

Hábito del zapallito amargo a mitad de temporada.
-

Zarcillos en los nudos de Cucurbita maxima.
-

Cucurbita maxima: Hoja con forma de riñón.
-

Cucurbita maxima: Hoja redonda.
-

Cucurbita maxima: Hoja con tres lóbulos.
-

Cucurbita maxima subsp. andreana: hoja con manchas.






Las flores son similares a las de los zapallos cultivados. En la misma planta hay flores masculinas y femeninas. Las masculinas son más numerosas y tienen un tallo largo, mientras que las femeninas son menos y tienen un tallo más corto y rígido. Las abejas y otros insectos son los encargados de llevar el polen de una flor a otra.
Los frutos del zapallito amargo se diferencian de los cultivados en varios aspectos:
- Tallos del fruto: En el zapallito amargo, el tallo que une el fruto a la planta es más blando y se cae fácilmente. En los zapallos cultivados, es más duro y permanece unido.
- Frutos maduros: Los frutos del zapallito amargo tienen muy poca pulpa, son secos y livianos, y se conservan por mucho tiempo. Los zapallos cultivados tienen pulpa jugosa, son pesados y se echan a perder más rápido.
- Sabor: Los frutos del zapallito amargo son amargos, mientras que los zapallos cultivados son dulces.
.jpg)
Las semillas son de color gris, lisas y brillantes, y miden entre 6 y 16 milímetros. Son mucho más livianas que las semillas de los zapallos cultivados y contienen una buena cantidad de grasas. También tienen más calcio que las semillas de los zapallos que comemos.
Las flores aparecen entre diciembre y abril y duran unas 9 horas. Son polinizadas principalmente por abejas.

Cuando el fruto del zapallito amargo madura, su cáscara se vuelve dura. Su sabor amargo se debe a unas sustancias llamadas cucurbitacinas, que son desagradables y tóxicas para la mayoría de los animales. Por eso, no hay muchos animales que se coman estos frutos y ayuden a dispersar sus semillas. Las semillas suelen germinar en el mismo lugar donde cayó el fruto. Sin embargo, algunos roedores, como las vizcachas, pueden llevarse las semillas para comerlas, y la actividad humana también puede ayudar a dispersarlas.
Algunas plantas de zapallito amargo tienen unas pequeñas estructuras en las hojas que se llaman nectarios extraflorales, pero no se sabe bien para qué sirven, ya que no se ven insectos visitándolos.
Las cucurbitacinas, que dan el sabor amargo, son sustancias que protegen a la planta de la mayoría de los animales que intentan comérsela. Sin embargo, algunos escarabajos, como la "vaquita" (Diabrotica speciosa), pueden detectarlas y hasta usarlas para su propia defensa.
¿Para qué se usa el zapallito amargo?
Como planta no deseada en cultivos

Los frutos amargos que a veces aparecen en los cultivos de zapallito son un problema. La ANMAT (Administración Nacional de Medicamentos, Alimentos y Tecnología Médica) describe estos frutos como pequeños, redondos y de cáscara dura, con muchas semillas y poca pulpa. Su consumo puede causar malestar estomacal y pérdida de líquidos, y cocinar el fruto no elimina este riesgo. Este problema se ha reportado en Argentina y Uruguay.
Como mencionamos, el polen del zapallito amargo puede llegar a las flores de los zapallos cultivados. Aunque el fruto que se cosecha de esa planta madre no será amargo, las semillas de ese fruto sí podrían producir plantas con frutos amargos en la siguiente generación. Por eso, se recomienda eliminar el zapallito amargo de los campos de cultivo para evitar que se cruce con las variedades dulces.
Como elemento decorativo

Se ha propuesto usar los frutos maduros del zapallito amargo como objetos decorativos, de forma similar a como se usan otras calabazas. Sin embargo, esto solo sería recomendable en lugares donde no haya cultivos de zapallos dulces cerca, para evitar la propagación del gen del sabor amargo.
Consumo humano
El zapallito amargo es muy amargo y puede causar problemas de salud si se come. Se sabe que en el pasado, algunas comunidades indígenas y rurales consumían el fruto después de un proceso para quitarle lo amargo, como asarlo entre cenizas calientes. Sin embargo, los experimentos modernos no han logrado eliminar el amargor de esta manera.
La ANMAT recomienda descartar cualquier zapallito que tenga sabor amargo y no intentar quitarle el amargor.
En la medicina
Las cucurbitacinas del zapallito amargo tienen un efecto muy fuerte en el cuerpo. Antiguamente se usaban como un laxante potente, pero debido a su toxicidad, su uso medicinal fue prohibido en 1943.
En trampas para insectos
Se ha descubierto que la pulpa de los zapallitos amargos atrae mucho a la "vaquita" (Diabrotica speciosa), un insecto que es una plaga para las hortalizas. Esto es curioso porque las cucurbitacinas son tóxicas para la mayoría de los insectos. Se ha propuesto usar los zapallitos amargos como cebo en trampas para controlar a estos insectos.
Otros usos
_color.jpg)
Antiguamente, los frutos del zapallito amargo se usaban como recipientes o vasijas.
En algunas regiones de Argentina, como San Luis, es una tradición recolectar los frutos del zapallito amargo, llamados "cháncaras", para los fogones de San Juan. Se colocan en el fuego y, al calentarse, explotan con un ruido fuerte, como si fueran fuegos artificiales.
También se ha encontrado que en Perú, algunas comunidades indígenas usan los frutos para hacer pequeñas artesanías que venden a los turistas.
¿Cómo se le llama al zapallito amargo?
Además de "zapallito amargo" o "zapallo amargo", esta planta tiene otros nombres, dependiendo de la región:
- Cháncara: en San Luis y Córdoba.
- Yeruá: en Entre Ríos y Uruguay.
- Zapallo chirigua
- Zapallo de vizcachera
- Vüre dapallu: en lengua ranquel, en La Pampa.
- Huevo de pato, zapallo pouaiquito, ishicopé, ishcopiqui: en Perú, por los indígenas.
- Calabazo o Porongo: nombres generales para recipientes hechos de calabazas, que a veces se aplicaban también a esta planta.
Galería de imágenes
-

Zapallito amargo, fotos de F. López Anido. Los 4 de la derecha son como los descriptos por Millán (1945) y otros autores.
-
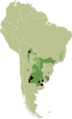
Mapa de distribución probable de zapallito amargo.
-
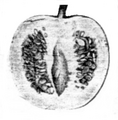
Fruto de la población de Jesús María (Córdoba, Argentina) en 1945. Corte transversal.
-

Fruto de la población de Jesús María (Córdoba, Argentina) en 1945. Fruto con corona y anillo.
-

Misma población, frutos sin (izquierda) y con (derecha) lóbulos.
-

Resultado de los experimentos de hibridación entre un zapallito cultivado y uno silvestre.
-

Semillas germinando en el lugar donde el fruto se formó.

.svg)





Véase también
 En inglés: Cucurbita andreana Facts for Kids
En inglés: Cucurbita andreana Facts for Kids
