
Red trófica para niños
Una red alimentaria (también conocida como red trófica o red de alimentos) es como un mapa que muestra quién se alimenta de quién en un ecosistema. Imagina todas las cadenas alimentarias de un lugar, como un bosque o un océano, conectadas entre sí; eso es una red alimentaria. Nos ayuda a entender cómo la energía y los nutrientes se mueven en la naturaleza.

Los científicos que estudian la ecología clasifican a los seres vivos en diferentes grupos, llamados niveles tróficos, según cómo obtienen su alimento. Estos grupos son:
- Productores: Son como las plantas, que fabrican su propio alimento.
- Consumidores primarios: Son los animales que comen plantas (herbívoros).
- Consumidores secundarios: Son los animales que comen a los herbívoros (carnívoros).
- Consumidores terciarios: Son los animales que comen a los consumidores secundarios.
- Descomponedores: Son organismos como hongos y bacterias que se encargan de reciclar la materia muerta.
Para crecer y vivir, los productores (como las plantas) crean su propia comida usando sustancias simples como minerales y dióxido de carbono. Para esto, necesitan energía, que casi siempre viene del Sol, a través de un proceso llamado fotosíntesis. Las conexiones en una red alimentaria muestran cómo los animales obtienen su energía al comer plantas o a otros animales.
La idea de las redes alimentarias fue desarrollada por científicos como Charles Elton en 1927. Antes, otros pensadores como Charles Darwin ya hablaban de la "red de la vida" para describir las complejas relaciones en la naturaleza.
Las redes alimentarias son una forma simplificada de representar los ecosistemas. Los científicos las usan para crear modelos y entender mejor cómo funcionan los sistemas donde unos organismos se alimentan de otros.
Contenido
Redes Alimentarias: ¿Quién Come a Quién en la Naturaleza?
Las conexiones en una red alimentaria nos muestran un mapa de las relaciones de alimentación en una comunidad de seres vivos. Los ecólogos dividen a todos los seres vivos en dos grandes grupos tróficos: los autótrofos y los heterótrofos.
Los autótrofos (productores) obtienen energía para vivir y crecer, ya sea del Sol (fotosíntesis) o de reacciones químicas. Los heterótrofos (consumidores) no producen su propio alimento; ellos comen a otros organismos para obtener la energía que necesitan. Una red alimentaria nos muestra cómo los heterótrofos se alimentan de los autótrofos y de otros heterótrofos, creando un flujo de energía y nutrientes.
El número de conexiones que tiene un consumidor en la red nos dice qué tan conectado está. Las cadenas alimentarias son como caminos rectos dentro de la gran red alimentaria, donde un organismo se come a otro en una secuencia.
Las conexiones en una red alimentaria unen "nodos", que son grupos de especies que tienen los mismos depredadores y presas. Por ejemplo, los parásitos, los microbios, los descomponedores, los consumidores y los depredadores son ejemplos de estos nodos.
¿Qué Son los Niveles Tróficos?

Las redes alimentarias tienen diferentes niveles o posiciones tróficas. Las especies que están en la base, como las plantas, forman el primer nivel. Ellas no se alimentan de ninguna otra criatura en la red. Estas especies basales pueden ser autótrofos o detritívoros (organismos que se alimentan de materia orgánica muerta). La mayoría de los autótrofos capturan la energía del Sol, pero algunos, como ciertas bacterias, obtienen energía de reacciones químicas en lugares oscuros, como las aguas termales.
En la cima de la red están los "superdepredadores", que son animales que ninguna otra especie puede cazar directamente para alimentarse. Los niveles intermedios están llenos de omnívoros, que comen de más de un nivel trófico, haciendo que la energía fluya en muchas direcciones.
En el esquema más sencillo, el primer nivel trófico son las plantas, el segundo son los herbívoros (que comen plantas) y el tercero son los carnívoros (que comen herbívoros). Los científicos estudian lo que comen los animales para clasificarlos en estos niveles. Hoy en día, se sabe que muchos organismos son omnívoros, lo que hace que las clasificaciones sean más complejas.
¿Cómo se Mueve la Energía en la Red Alimentaria?
El concepto de nivel trófico fue muy importante para entender cómo se transfiere la energía de una parte del ecosistema a otra. Aunque los omnívoros no se limitan a un solo nivel, los estudios recientes confirman que los niveles tróficos sí existen.
Un tema importante en la ecología es cómo se controlan las poblaciones de los seres vivos. Por ejemplo, los herbívoros a menudo tienen mucha comida (plantas), lo que significa que sus poblaciones son controladas principalmente por sus depredadores. Esto se conoce como la hipótesis de "arriba-abajo". Pero también es cierto que no toda la materia vegetal se consume, y la calidad de las plantas puede influir en los herbívoros, lo que sugiere un control de "abajo-arriba". Los estudios muestran que ambos tipos de fuerzas pueden influir en un ecosistema.
Otro ejemplo de estas interacciones es la "cascada trófica". Aquí, los depredadores pueden ayudar a que las plantas crezcan más al evitar que los herbívoros las coman en exceso. Aunque los depredadores no comen plantas directamente, controlan la población de los herbívoros que sí lo hacen. Este efecto indirecto es una cascada trófica.
Flujo de Energía y Biomasa
Las redes alimentarias nos muestran cómo la energía se mueve a través de las conexiones tróficas. El flujo de energía va en una sola dirección, a diferencia de la materia, que se recicla. La energía se usa para crecer, moverse y respirar, y una parte se pierde en cada paso.
La masa de un organismo (su biomasa) está relacionada con su contenido de energía. La energía se "degrada" y se pierde en forma de calor a medida que pasa de un nivel trófico a otro. Por eso, la biomasa de cada nivel trófico disminuye desde la base de la cadena hacia la cima.
Aproximadamente del 80% al 90% de la energía se pierde como calor o desechos en cada transferencia. Solo entre el 10% y el 20% de la energía pasa al siguiente organismo. Las representaciones gráficas de la biomasa o la productividad en cada nivel se llaman pirámides ecológicas o tróficas.
Pirámides Ecológicas



En una pirámide de números, la cantidad de consumidores disminuye mucho en cada nivel. Por ejemplo, un solo superdepredador (como un oso polar) necesita una cantidad mucho mayor de productores para sobrevivir. Generalmente, hay un máximo de cuatro o cinco conexiones en una cadena alimentaria. Al final, toda la energía en una cadena se convierte en calor.
Las pirámides ecológicas muestran a los productores primarios en la base. Representan diferentes características de los ecosistemas, como el número de individuos, la biomasa (peso seco de los organismos) y la energía. La forma de pirámide, con menos energía en los niveles superiores, es un patrón que se repite en casi todos los ecosistemas del planeta.
A veces, las pirámides de biomasa pueden estar "invertidas", especialmente en ecosistemas acuáticos como los arrecifes de coral. Esto ocurre porque los productores acuáticos, como las algas, son muy pequeños y se reproducen muy rápido, lo que les permite soportar una biomasa más grande de consumidores. Sin embargo, las pirámides de energía siempre tienen una forma normal, con la base más ancha, porque la energía siempre se pierde en cada transferencia, siguiendo las leyes de la física.
Flujo de Materia y Reciclaje
Muchos de los elementos y minerales de la Tierra se encuentran dentro de los seres vivos. Por eso, los ciclos de minerales y nutrientes siguen las mismas rutas que el flujo de energía en las redes alimentarias. Los ecólogos estudian elementos clave como el carbono (C), el nitrógeno (N) y el fósforo (P).
Los nutrientes minerales son esenciales para que los organismos crezcan y se desarrollen. Las redes alimentarias describen cómo estos minerales fluyen a través de los organismos. Gran parte de la materia producida en un ecosistema no es consumida directamente, sino que es reciclada por los descomponedores (como bacterias y hongos) y vuelve a ser útil para las plantas.
Tipos de Redes Alimentarias
Las redes alimentarias son una forma simplificada de mostrar la complejidad de los ecosistemas reales. Hay muchísimas especies en el planeta, y la mayoría son microbios e invertebrados. Los científicos se enfocan en las interacciones más importantes, donde ocurre la mayor parte de la transferencia de energía.
Existen diferentes tipos de redes alimentarias, según lo que se quiera estudiar:
- Red fuente: Muestra un organismo y todo lo que se alimenta de él, y así sucesivamente.
- Red hundida: Muestra un organismo y todo lo que él come, y lo que comen sus presas.
- Red de comunidad: Muestra un grupo de organismos y todas las conexiones de quién come a quién.
- Red de flujo de energía: Mide la cantidad de energía que se mueve entre los organismos.
- Red paleoecológica: Reconstruye ecosistemas antiguos a partir de fósiles.
- Red funcional: Destaca las conexiones más importantes que tienen un mayor impacto en la organización de la comunidad.
Dentro de estas categorías, las redes alimentarias se pueden organizar según el tipo de ecosistema, como redes alimentarias marinas, terrestres, del suelo, o incluso las que involucran a los humanos.
Red Detrítica
En una red detrítica, la materia muerta de plantas y animales es descompuesta por bacterias y hongos. Luego, esta materia es consumida por detritívoros (como gusanos) y, a su vez, estos son comidos por carnívoros. A menudo, hay conexiones entre la red detrítica y la red de "pastoreo" (donde los herbívoros comen plantas vivas). Por ejemplo, los hongos que crecen de la materia descompuesta pueden ser alimento para ciervos o ardillas.

El detrito es cualquier tipo de materia orgánica no viva, como hojas caídas, madera muerta, restos de animales o heces. La importancia de cada tipo de detrito varía según el ecosistema.
Redes Alimentarias Cuantitativas
Los ecólogos recogen datos sobre los niveles tróficos y las redes alimentarias para crear modelos matemáticos. Esto les permite estudiar patrones y propiedades que se repiten en diferentes ecosistemas. Una red alimentaria puede mostrar cómo un cambio en una sola especie puede afectar a muchas otras, tanto directa como indirectamente.
Aunque la complejidad de las redes alimentarias reales es difícil de entender por completo, los modelos matemáticos son herramientas muy valiosas. Nos ayudan a comprender la estructura, la estabilidad y las reglas que rigen el comportamiento de estas redes.
La complejidad de una red alimentaria se mide por el número de especies y sus conexiones. Se pensaba que una mayor complejidad llevaba a una mayor estabilidad en las redes alimentarias, y estudios recientes han confirmado que esto es cierto en los sistemas reales.
Las redes alimentarias son complejas porque cambian de tamaño según las estaciones o la geografía. Los organismos y los nutrientes minerales pueden moverse entre diferentes ecosistemas.
Historia de las Redes Alimentarias

Las redes alimentarias son una herramienta fundamental para los ecólogos, que les permite organizar las complejas interacciones entre las especies en la naturaleza. Una de las primeras descripciones de una cadena alimentaria fue hecha por el académico medieval Al-Jahiz, quien dijo: "Todos los animales, en resumen, no pueden existir sin comida, ni tampoco puede el animal cazador escapar a ser cazado".
Las primeras representaciones gráficas de una red alimentaria aparecieron a finales del siglo XIX y principios del XX. Charles Elton fue un pionero en el concepto de las redes alimentarias en su libro de 1927, "Ecología Animal". Después de su trabajo, las redes alimentarias se convirtieron en un concepto central en el estudio de la ecología.
El interés en las redes alimentarias creció aún más después de los estudios de Robert Paine, que sugirieron que la complejidad de estas redes era clave para mantener la diversidad de especies y la estabilidad de los ecosistemas. Esto llevó a muchos ecólogos a estudiar las propiedades matemáticas de las redes alimentarias.
Galería de imágenes
-
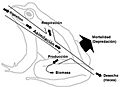
Diagrama de flujo de energía de una rana. La energía ingerida se usa para vivir y se transforma en biomasa.
-
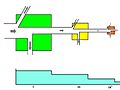
Una cadena alimentaria de tres partes que muestra cómo la energía se "degrada" y pierde calidad a lo largo de la cadena.


Véase también
 En inglés: Food web Facts for Kids
En inglés: Food web Facts for Kids
